弥漫大B细胞淋巴瘤研究中的FAS脂肪酸合成酶活性检测——脂代谢重编程如何成为治疗靶点
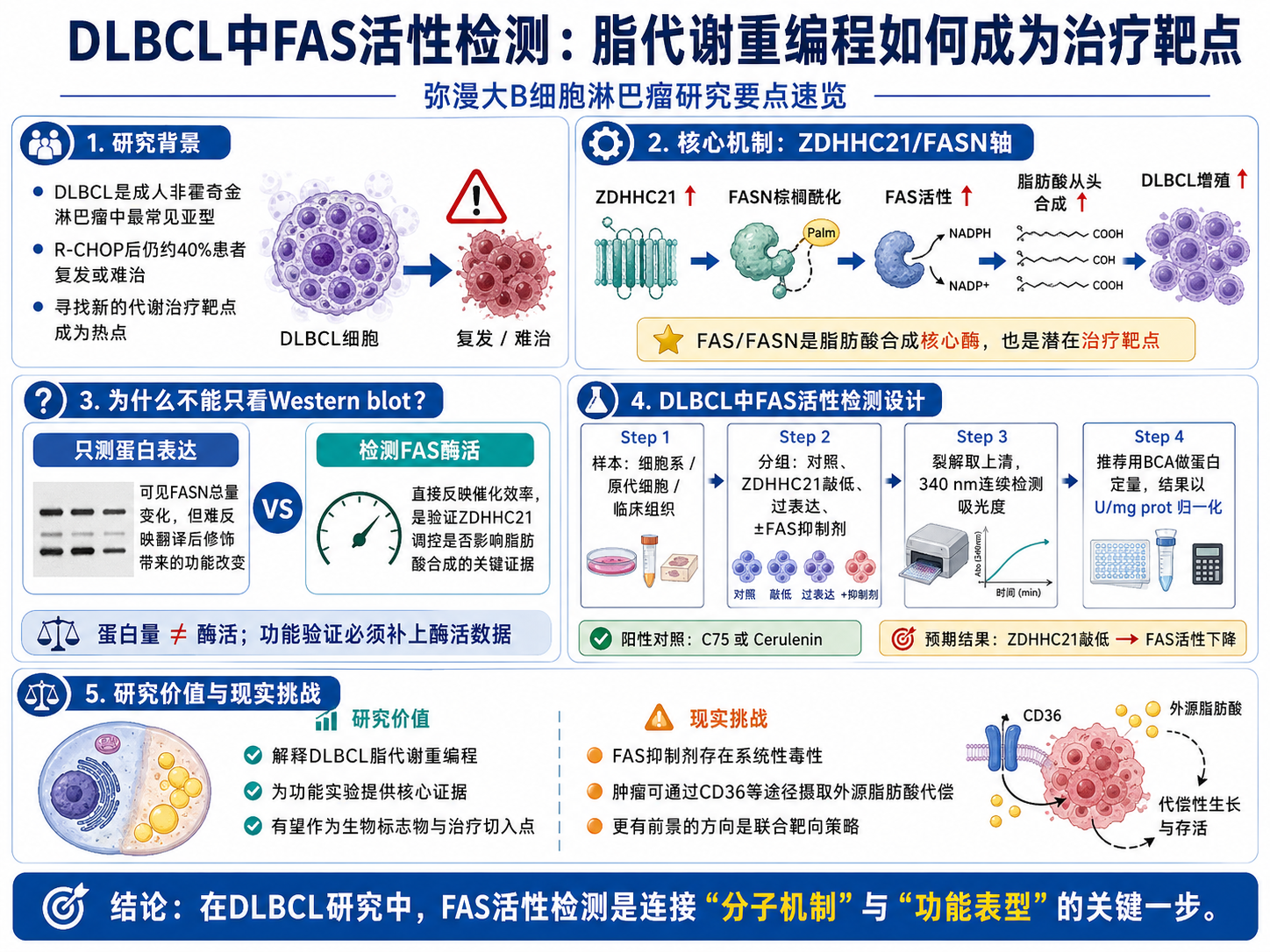
弥漫大B细胞淋巴瘤(Diffuse Large B-Cell Lymphoma,DLBCL)是成人非霍奇金淋巴瘤中发病率最高的亚型,占所有病例的30%到40%。尽管R-CHOP方案(利妥昔单抗联合化疗)已经成为标准一线治疗,仍有约40%的患者会在治疗后复发或出现难治性病变。这批患者的预后很差,现有的挽救治疗方案有效率有限,寻找新的治疗靶点一直是DLBCL研究领域的核心任务之一。
过去十年里,肿瘤代谢重编程作为一个治疗靶点方向获得了越来越多的关注。DLBCL细胞的增殖速度极快,对能量和生物合成原料的需求远超正常B细胞,这种高代谢需求必须依赖特定的代谢通路来支撑。脂肪酸的从头合成,正是其中一条被反复证实在DLBCL中异常激活的代谢通路,而FAS(脂肪酸合成酶)就是这条通路的核心执行者。
肿瘤细胞为什么需要大量合成脂肪酸
正常成体细胞在绝大多数情况下不依赖内源性脂肪酸合成,它们更倾向于直接从血液中摄取现成的脂肪酸来满足自身的膜脂更新和能量需求。这是一种能量上非常经济的策略——不需要自己造,从外面取就行。
肿瘤细胞的情况完全不同。快速增殖需要大量的磷脂来构建新的细胞膜,需要脂肪酸作为信号分子的前体,也需要脂质参与蛋白质的翻译后修饰。仅靠外源摄取往往无法满足这种急剧扩张的需求,于是肿瘤细胞重新激活了在正常成体组织中几乎处于沉默状态的内源性脂肪酸合成程序。FAS作为这个程序的核心酶,在多种肿瘤中都出现了表达量上调和活性增强的现象,包括乳腺癌、前列腺癌、结直肠癌,以及近年来越来越受到关注的血液肿瘤——DLBCL。
FAS在肿瘤中的这种角色,让它天然地成为了一个潜在的治疗靶点:如果能够抑制肿瘤细胞的FAS活性,从源头上切断脂肪酸的供应,是否能够遏制肿瘤细胞的增殖?这个逻辑在实验室层面已经得到了反复验证,FAS抑制剂在多种肿瘤模型中表现出了肿瘤抑制效应,但如何在临床上实现精准的靶向干预,仍然需要更深入地理解FAS在特定肿瘤类型里的调控机制。
ZDHHC21/FASN轴:一个在DLBCL中被发现的新调控节点
2023年发表于Leukemia的研究,把视角聚焦在了一个之前在DLBCL背景下研究不多的蛋白——ZDHHC21。
ZDHHC21是一种棕榈酰转移酶,负责催化蛋白质的S-棕榈酰化修饰,也就是把棕榈酸共价连接到蛋白质的半胱氨酸残基上。棕榈酰化是一种可逆的脂质修饰,能够调控靶蛋白的膜定位、稳定性和蛋白质相互作用,在细胞信号传导中发挥重要作用。
这项研究发现,ZDHHC21在DLBCL中表达上调,而它的一个重要靶蛋白,正是FASN——也就是FAS的编码蛋白。ZDHHC21通过棕榈酰化修饰FASN,影响其稳定性和活性,从而在上游调控DLBCL细胞的脂肪酸合成能力。换句话说,ZDHHC21并不直接参与脂肪酸合成,但它通过对FAS蛋白的修饰,间接控制了整个合成通路的输出。
这个发现的意义在于,它揭示了一条新的调控轴:ZDHHC21→FASN→脂肪酸合成→DLBCL增殖。如果能够靶向ZDHHC21,就有可能在不直接作用于FAS的情况下,间接抑制DLBCL细胞的脂肪酸合成,为DLBCL的治疗提供一个新的干预点。
在这个研究框架里,FAS活性检测承担了一个关键的功能验证角色——研究者需要证明的不只是ZDHHC21影响了FASN的蛋白表达量或者稳定性,而是这种影响真实地改变了FAS的催化效率,从功能层面改变了细胞的脂肪酸合成能力。这个区别很重要:蛋白量的变化是上游事件,酶活的变化才是下游功能性结果,两者加在一起,证据链才是完整的。
为什么单靠Western blot不够
做淋巴瘤相关的FAS研究,很多研究者的第一反应是跑一个Western blot,看FASN蛋白条带的深浅,再配合qPCR看一下mRNA水平。这是标准的起步操作,本身没有问题,但在ZDHHC21/FASN这个具体的研究场景里,它有一个根本性的局限。
ZDHHC21对FASN的调控,主要发生在翻译后修饰层面——棕榈酰化影响的是蛋白的构象、定位和稳定性,不一定会在总蛋白量上产生显著变化。如果ZDHHC21的敲低或过表达让FASN的棕榈酰化水平改变了,但FASN的总蛋白量变化不大,Western blot就会告诉你"没什么差别"——而实际上,FAS的催化活性可能已经发生了显著变化。
这种情况在翻译后修饰研究里非常常见。修饰改变了酶的构象,影响了活性位点的可及性,或者改变了酶与辅因子的结合效率,最终体现在催化速率的变化上,但在总蛋白量上完全看不出来。这就是为什么在这类研究里,酶活检测是不可替代的——它直接测量的是FAS"在干多少活",而不是"有多少FAS蛋白在那里"。
从审稿的角度来看,如果一篇文章声称某种调控影响了FAS功能,却只有蛋白量数据而没有活性数据,审稿人很可能会要求补充酶活实验。Leukemia这样的高分期刊,对功能验证数据的完整性要求尤其严格。FAS活性数据在这里不是锦上添花,是必要的功能性证据。
DLBCL研究中FAS活性检测的样本类型和实验设计
在DLBCL的研究场景里,用于FAS活性检测的样本类型主要有三类:细胞系、原代细胞和临床样本。每一类的处理方式和数据解读逻辑都有所不同,需要分开来说。
细胞系是最常见的起点。 OCI-LY3、OCI-LY10、SUDHL-4、SUDHL-6等都是DLBCL研究里常用的细胞系,覆盖了ABC亚型和GCB亚型。用这些细胞系做FAS活性检测,前处理相对标准:收集5×10⁶个细胞,预冷PBS清洗去除培养基,800 g离心2分钟弃上清,加入1 mL Extraction Buffer进行冰浴超声裂解(功率20%或200 W,超声3秒、间隔7秒、重复30次),之后12000 g、4℃离心40分钟取上清。
淋巴瘤细胞系大多是悬浮培养,收集比贴壁细胞方便,不需要胰酶消化这个步骤,但离心收集的时候要确保细胞沉淀完全,上清吸干净再加裂解液,否则残留的培养基会稀释样本,影响后续的蛋白浓度测定和酶活计算。
在ZDHHC21/FASN的研究框架下,细胞系实验通常需要设计多个对比组:ZDHHC21正常表达组(对照)、ZDHHC21敲低组(siRNA或shRNA)、ZDHHC21过表达组,有时候还需要加入FAS抑制剂(如C75或Cerulenin)作为阳性对照,验证观察到的FAS活性变化确实来自FAS本身而不是其他干扰因素。这些组的FAS活性数据放在一起,才能完整展示ZDHHC21对FAS活性的调控效应。
原代细胞的情况更复杂一些。 从DLBCL患者外周血或者淋巴结中分离的原代肿瘤细胞,在生物学特性上比细胞系更接近真实的疾病状态,但数量往往更有限,而且细胞活力在分离过程中可能受到影响。处理方式和细胞系基本一致,但需要特别注意的是,原代细胞通常比细胞系更脆弱,超声裂解的参数可能需要适当调整,确保裂解充分的同时避免过度破坏导致样本损失。
如果原代细胞数量不够5×10⁶,可以适当减少,但需要在上样体积上做对应调整,或者在数据解读时对蛋白归一化的结果保持谨慎。当细胞数量过少,单个样本的ΔA值本来就很小,这时候数据的信噪比会下降,结果的可重复性也会变差。
临床组织样本。 淋巴结活检组织是另一类重要的样本来源。取约0.1 g组织,加入1 mL Extraction Buffer,在冰浴条件下充分匀浆,12000 g、4℃离心40分钟取上清。淋巴结组织的质地比实质性器官(比如肝脏)松软,匀浆通常比较容易,但活检样本来之不易,操作前必须确认试剂和仪器都已准备就绪,不能因为操作失误浪费样本。
检测流程中值得特别注意的几个环节
DLBCL研究场景有一些和普通组织检测不太一样的地方,在操作上需要格外注意。
淋巴瘤细胞是高代谢状态的细胞,FAS活性通常比正常静止期细胞高出不少。这意味着ΔA值可能相对较高,在ΔA超过0.4的情况下需要稀释样本重新检测。正式大批量实验之前做一轮预实验,摸清楚自己手头细胞系或者原代细胞的大致ΔA范围,可以避免整批数据因为超出线性范围而报废。
工作液的配制和预孵育同样不能省。每孔180 µL工作液的组成是:16 µL Working NADPH、4 µL Working Acetyl CoA、8 µL Working Malonyl CoA、152 µL Assay Buffer,配好之后在37℃孵育15分钟以上再开始加样。在比较多个处理组的实验里,如果工作液没有充分预孵育,不同孔之间反应启动时间不一致,会在组间引入系统误差,让本来应该有差异的组看起来没什么差别,或者让正常的数据波动被误读为真实差异。
在多组对比的实验设计里,建议同一批实验里所有组的细胞在相同时间点收集,裂解液在同一天制备,FAS活性检测当天完成。不同批次制备的裂解液如果需要跨天比较,批次间的变异会给数据带来额外的不确定性,结果的解读会变得复杂。
数据归一化:在淋巴瘤研究里这一步特别重要
做肿瘤细胞的FAS活性检测,归一化方式的选择比做组织样本时更需要认真对待。
在ZDHHC21敲低或过表达的实验里,处理本身可能影响细胞的增殖速率和存活率,导致不同处理组在收集时的实际细胞状态存在差异。如果直接按细胞数量归一化(U/10⁴细胞),但各组细胞的平均蛋白含量因为处理不同而有差异,归一化结果就不能消除这种差异。
这种情况下,推荐在收集细胞做FAS活性检测的同时,用同一批裂解液做蛋白定量,以蛋白浓度归一化(U/mg prot)作为主要的结果表述方式。蛋白浓度归一化能够消除由于细胞增殖状态差异带来的影响,让不同处理组之间的FAS活性具有真正可比较的基础。
用于蛋白定量的BCA法是目前最常用的选择,对裂解液中常见的去垢剂有一定耐受性,和Extraction Buffer的兼容性通常没有问题。亚科因生物的KTB2240和配套的BCA蛋白定量试剂盒(KTD3001)可以用同一批上清同步检测,减少因为重复取样带来的操作误差。
FAS活性数据在DLBCL研究里如何解读
拿到FAS活性数值之后,怎么解读这个数据,在DLBCL研究里有一些特定的逻辑需要梳理清楚。
如果比较的是正常B细胞和DLBCL细胞: DLBCL细胞的FAS活性通常显著高于静止期的正常B细胞,这和文献里对肿瘤细胞FAS高表达的普遍描述一致。如果你自己的数据里DLBCL细胞系的FAS活性并不比正常B细胞高,需要先排查操作问题,比如正常B细胞的激活状态——被激活的正常B细胞(比如经过抗原刺激的生发中心B细胞)FAS活性也会显著升高,如果用激活状态的正常B细胞做对照,两者的差异会缩小。
如果比较的是ZDHHC21敲低前后的DLBCL细胞: 根据Leukemia文章的研究逻辑,ZDHHC21敲低预期会降低FAS活性,因为失去了ZDHHC21的棕榈酰化支持,FASN蛋白的稳定性或活性构象可能受到影响。如果FAS活性在敲低后显著下降,配合Western blot里FASN蛋白量的变化(或者不变化),就能支撑"ZDHHC21通过调控FASN功能状态影响脂肪酸合成"这个结论。如果FAS活性没有变化,则需要考虑调控是否发生在其他层面,或者是否有补偿机制存在。
如果配合脂肪酸合成抑制剂使用: 加入C75(FAS抑制剂)或者Cerulenin(FAS特异性抑制剂)的处理组,FAS活性应该显著下降,接近空白值。这个数据一方面是阳性对照,验证检测体系本身是有效的;另一方面,抑制剂处理后FAS活性下降的幅度,结合细胞增殖和凋亡数据,能够定量地评估FAS活性对DLBCL细胞存活的贡献程度。
这个研究方向目前的进展和值得关注的问题
DLBCL的脂代谢研究这几年进展很快,但也有一些尚待解决的问题,以及一些需要客观面对的现实局限,对于正在或者计划进入这个方向的研究者来说,了解这些背景有助于设计更有针对性的实验。
DLBCL并不是一个均一的疾病,按照细胞起源可以分为ABC亚型(活化B细胞样)和GCB亚型(生发中心B细胞样),两种亚型在代谢表型上存在差异。目前对于FAS在两种亚型中的活性差异,以及ZDHHC21/FASN轴的调控机制是否在两种亚型中都同样适用,还没有非常系统的数据。如果你的研究同时涵盖了ABC和GCB亚型的细胞系,同时报告两种亚型的FAS活性数据,会让结果更有说服力,也更符合近年来DLBCL研究对亚型差异的重视程度。
关于FAS作为治疗靶点,有一个现实必须说清楚。
FAS抑制剂在细胞系和动物模型里的抗肿瘤效果,已经被反复验证,这一点是真实的。但C75和Cerulenin这两个在实验室里最常用的FAS抑制剂,在临床前研究里暴露出了非常严重的毒副作用——剧烈的体重下降、厌食、以及神经毒性。这些不是可以通过剂量优化轻易解决的边缘问题,而是FAS在正常能量代谢中的生理角色所决定的系统性毒性。FAS在肝脏、脑等正常组织中同样高表达,全身性抑制FAS势必影响正常组织的脂肪酸合成,这是几十年来FAS抑制剂始终难以真正进入临床获批的核心障碍。
所以当我们说ZDHHC21/FASN轴是一个"潜在的治疗靶点",这个"潜在"是有前提的——从机制研究的价值来说,揭示这条调控轴无疑是重要的;但要从靶向ZDHHC21或FASN发展出临床可用的治疗策略,中间还有很长的路。这并不否定研究的价值,但需要对临床转化的预期保持清醒。
另一个需要认真对待的问题是代谢代偿。
"抑制FAS就能切断肿瘤细胞的脂肪酸供应"这个逻辑,在体外实验里往往成立,但在体内环境里是被简化了的。真实生理环境里,DLBCL细胞在FAS活性被抑制之后,通常不会坐以待毙——它们会迅速上调细胞膜上的脂质转运蛋白(比如CD36),转而大量摄取血液中的外源性游离脂肪酸来代偿内源合成的不足。这种代偿策略的启动速度很快,是肿瘤细胞代谢可塑性的典型体现。
这也直接解释了一个在体内实验里反复出现的现象:在细胞系里效果很好的FAS靶向策略,到了动物模型里,或者在有外源脂质供给的体内环境里,效果会大打折扣,耐药出现的也比预期快。单一靶向ZDHHC21或FASN,如果不同时阻断外源脂肪酸的摄取通路,在体内很可能是不够的。
这不是说这个方向没有价值,而是说更有前景的策略可能是组合靶向:同时抑制内源合成(FAS)和外源摄取(CD36等转运蛋白),或者把FAS靶向和其他代谢干预手段结合起来。这是目前肿瘤脂代谢领域正在积极探索的方向,也是DLBCL代谢研究下一步最值得关注的问题之一。
另外,FAS活性作为生物标志物,在DLBCL的精准治疗语境下也有潜在价值。如果某类患者的DLBCL细胞表现出特别高的FAS活性,这些患者是否会对特定代谢干预策略有更好的响应?这个假说目前还在研究层面,但FAS活性检测是验证这个假说所需的基础技术手段。在有临床随访数据的患者样本中同时检测FAS活性,探索其与治疗反应或生存期的相关性,是一个兼具科学价值和临床转化意义的研究方向。
在论文方法部分如何描述
对于使用KTB2240(亚科因生物,CheKine™脂肪酸合成酶活性检测试剂盒,微量法)做FAS活性检测的研究,在方法部分可以参考以下描述方式:
FAS活性检测按照试剂盒说明书(CheKine™ Micro Fatty Acid Synthetase Activity Assay Kit,KTB2240,Abbkine Scientific)进行。简述如下:收集细胞后用预冷PBS清洗,加入Extraction Buffer超声裂解,12000 g、4℃离心40分钟取上清。每孔检测体系为20 µL样本上清与180 µL工作液(含NADPH、乙酰CoA、丙二酰CoA)的混合,在96孔UV板中于37℃孵育后,在酶标仪上连续检测340 nm处吸光度。记录第10秒(A₁)和第70秒(A₂)的吸光度值,以ΔA = A₁ - A₂计算FAS活性,结果以U/mg蛋白表示,蛋白浓度使用BCA法测定。
这种描述方式引用了试剂盒的商品名、货号和制造商,同时给出了足够的操作细节,满足方法部分的可重复性要求,也为审稿人提供了验证方法可靠性的引用依据。
为什么DLBCL是FAS活性研究最值得投入的方向之一
如果你的课题在血液肿瘤方向,FAS活性检测是一个回报率非常高的实验投入。
原因是多方面的。首先,脂代谢重编程在DLBCL里有越来越强的文献支撑,这个方向的接受度在审稿层面已经建立,不需要花大量篇幅去说服审稿人为什么脂代谢在淋巴瘤里重要。其次,ZDHHC21/FASN这个调控轴是一个相对新的发现,下游的机制问题还有很多没有解答,围绕这个轴做延伸研究有充足的空间。第三,FAS活性检测本身操作不复杂,数据产出周期短,作为功能验证实验可以相对快速地为一篇文章补充必要的代谢表型证据。
更实际的考量是:Leukemia发表了ZDHHC21/FASN轴的研究,意味着这个期刊的编辑和审稿人对这个方向是认可的,后续在相关方向上做出新发现的稿件,在期刊选择上有了一个明确的参照点。而FAS活性数据,几乎必然会出现在这类投稿的核心数据里。
写在最后
DLBCL的研究者越来越多地意识到,理解这种疾病不能只停留在基因突变和信号通路,代谢层面的改变既是肿瘤细胞维持高速增殖的基础,也是潜在的治疗弱点。FAS作为脂肪酸从头合成的核心酶,在DLBCL里的角色正在从一个"有趣的现象"转变为一个"值得认真对待的靶点"。
检测FAS活性,是把这个靶点从蛋白表达层面落实到功能层面的关键一步。数据本身不复杂,操作也不难,难的是知道什么时候需要这个数据,以及怎么把它放进正确的实验设计框架里,让它在证据链里发挥应有的价值。
希望这篇文章对正在做DLBCL脂代谢相关研究的你有所帮助。
