Rubisco是什么?图解二磷酸核酮糖羧化酶/加氧酶的催化原理与活性检测逻辑

提到光合作用,大多数人脑子里浮现的是叶绿素、光反应、ATP。但有一个酶,既不发光、也不传递电子,却是整个光合作用能否真正"固碳"的最终关卡——它叫Rubisco,全称1,5-二磷酸核酮糖羧化/加氧酶(Ribulose-1,5-bisphosphate carboxylase/oxygenase,EC 4.1.1.39)。
如果你最近开始接触植物生理实验,或者课题里需要检测光合能力,Rubisco活性几乎是绕不开的一个指标。这篇文章从头讲清楚:Rubisco是什么、它做了什么、为什么连"最慢的酶"也不可替代,以及实验室里最常用的活性检测方法背后的逻辑是什么。
Rubisco是地球上含量最多的蛋白质,却也是最"慢"的关键酶
先说一个反直觉的事实:Rubisco是地球上含量最高的蛋白质,估计全球总量约7亿吨,占到植物叶片可溶性蛋白的50%左右。但与此同时,它的催化效率极低——催化转换数(kcat)仅约3–10 s⁻¹,意思是每个活性位点每秒只能催化3到10个CO₂分子完成固定反应。相比之下,大多数代谢酶的kcat在数百到数千之间,碳酸酐酶甚至高达10⁶ s⁻¹。
植物的"解决方案"是堆量:既然单个酶分子跑得慢,就拼命多合成一些,用数量弥补速度。这就是为什么一片叶子里Rubisco含量如此之高,也是为什么氮素是限制植物生产力的核心资源之一——大量氮被"锁"在了Rubisco分子里。
在气候变化和粮食安全的研究背景下,Rubisco的改良是近年来植物科学最热的方向之一:能不能提高它的催化速率?能不能增强它对CO₂的特异性、减少"走错方向"的概率?这些问题的回答,都需要从准确测定Rubisco的活性开始。
Rubisco做了什么:Calvin循环的入口反应
光合作用分两个阶段。光反应在类囊体膜上进行,把光能转化为ATP和NADPH;暗反应(Calvin循环)在叶绿体基质中进行,用这些能量把CO₂"固定"成有机物。
Rubisco催化的是Calvin循环的第一步,也是整个循环的核心限速步骤:
1分子RuBP(核酮糖-1,5-二磷酸)+ 1分子CO₂ → 2分子PGA(3-磷酸甘油酸)
这一步反应把无机碳(CO₂)并入有机分子骨架,是地球上几乎所有有机碳的最终来源。PGA随后经过一系列酶促反应,一部分用于合成糖、淀粉、氨基酸等有机物,另一部分用于再生RuBP,维持循环的持续运转。
没有这一步,光反应产生的ATP和NADPH就没有用武之地,光合作用等于前功尽弃。这就是Rubisco"关键酶"称号的由来——它是光能转化为化学能这条流水线的最终出口。
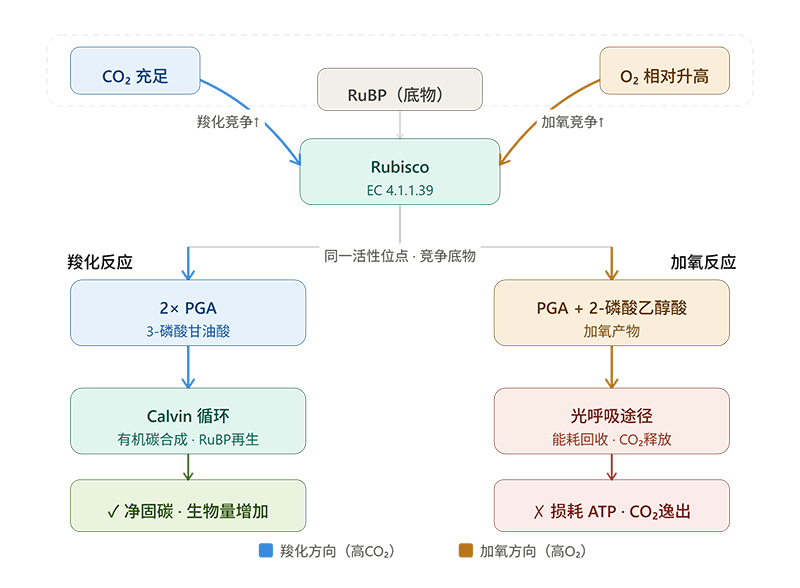
Rubisco的两面性:固碳还是"浪费碳",取决于CO₂/O₂的比值
Rubisco的麻烦在于它不"专一"。除了催化CO₂固定(羧化反应),它还会催化O₂与RuBP的反应(加氧反应),走向光呼吸途径:
1分子RuBP + 1分子O₂ → 1分子PGA + 1分子磷酸乙醇酸(2-PG)
磷酸乙醇酸需要经过一系列消耗能量的步骤才能被回收,这个过程会释放已经固定的CO₂,并消耗ATP和还原力——本质上是在"倒退",通常被称为光呼吸损耗。
羧化还是加氧,取决于活性位点周围CO₂和O₂的浓度比。叶绿体基质中CO₂浓度高时,羧化占优;O₂浓度相对升高时(比如气孔关闭导致CO₂供应减少、或高温导致CO₂溶解度下降),加氧比例上升,光呼吸损耗加剧,光合效率下降。
这正是为什么干旱和高温对C3植物(水稻、小麦、大豆等)的光合作用打击尤其严重:气孔关闭 → 叶肉细胞CO₂浓度下降 → Rubisco加氧反应比例上升 → 净固碳量下滑。Rubisco活性检测,正是量化这种打击程度的核心工具之一。
C4植物(玉米、甘蔗)和CAM植物通过特殊的碳浓缩机制,在Rubisco周围维持较高的CO₂浓度,有效抑制加氧反应,因此在高温干旱环境下光合效率更高。这一机制差异,也是C3/C4植物比较研究中Rubisco活性检测的重要背景。
实验室里如何检测Rubisco活性:NADH偶联比色法的逻辑
Rubisco的催化产物PGA本身无色,CO₂的消耗量也难以直接测量,因此商品化试剂盒普遍采用偶联酶法,把Rubisco催化反应的产物信号"传递"到一个可以光学检测的分子上。
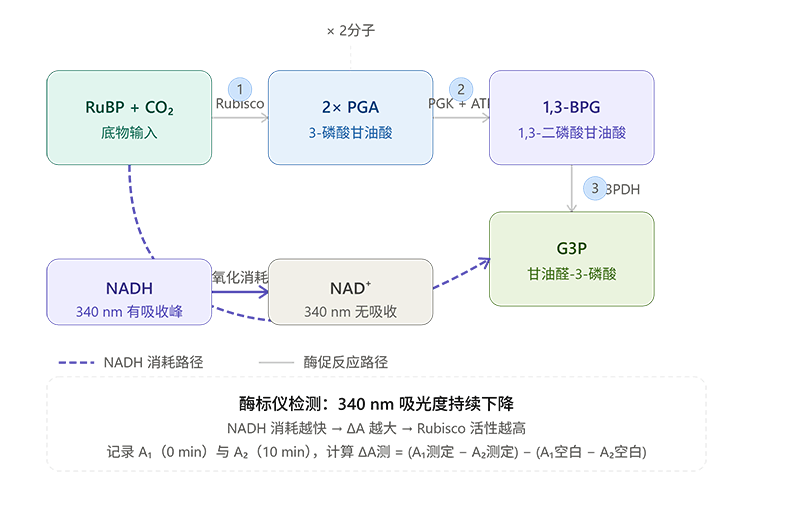
以CheKine™ 二磷酸核酮糖羧化酶/加氧酶(Rubisco)检测试剂盒(微量法)KTB1480为代表的NADH偶联比色法,完整信号链如下:
RuBP + CO₂ --[Rubisco]--> 2×PGA PGA + ATP --[PGK]--> 1,3-二磷酸甘油酸(1,3-BPG)+ ADP 1,3-BPG + NADH --[G3PDH]--> 甘油醛-3-磷酸(G3P)+ NAD⁺ + Pi
其中PGK是3-磷酸甘油酸激酶,G3PDH是甘油醛-3-磷酸脱氢酶,两者均由试剂盒预置提供。整个偶联链的净效果是:Rubisco每固定1分子CO₂,最终消耗2分子NADH。
NADH在340 nm有特征吸收峰,NAD⁺在该波长无吸收。因此,在96孔UV板上用酶标仪持续监测340 nm吸光度,吸光度的下降速率直接反映NADH的消耗速率,也就是Rubisco的羧化酶活性。
简单来说:NADH是这个检测体系里的"报告分子",它消失得越快,说明Rubisco越活跃。
反应体系总体积为0.21 mL(20 µL样本 + 190 µL工作液),在25℃下反应10 min,分别记录0 min(A₁)和10 min(A₂)时的吸光值,计算扣除空白后的ΔA,代入公式即可得到以U/g或U/mg蛋白表示的Rubisco活性。
这个方法测的是什么,不测的是什么
理解方法的边界,和理解方法本身一样重要。
测到的: 样本提取液在标准反应体系中的羧化酶活力,反映的是提取到反应体系里的Rubisco总量与其当下的催化能力之积。适合在不同处理组之间做相对比较。
没有测到的:
第一,加氧酶活性。本方法专门检测羧化反应,加氧反应(光呼吸方向)不在检测范围内。如需研究羧化/加氧比值,需要配合其他方法(如气体交换法或同位素示踪)。
第二,原位活化状态。Rubisco在细胞内存在"活化"和"失活"两种状态,受Rubisco活化酶(RCA)的调控,并随光照、温度、CO₂浓度实时变化。提取过程会打破这种动态平衡,标准单次检测得到的是"提取后的综合活力",不能直接等同于叶片原位的活化状态。若需区分初始活性和总活性,需设置额外的活化剂处理组(如碳酸氢钠/MgCl₂预处理),分别测定后计算活化百分比。
第三,Rubisco蛋白含量。酶活力≠蛋白量。两个处理组的Rubisco活性差异,可能来自蛋白量的变化,也可能来自活化状态的变化,或两者兼有。如需区分,需要配合Western blot或ELISA定量Rubisco蛋白量。
结果如何换算成活性单位
KTB1480提供两种归一化方式,对应两种单位定义:
按样本质量(U/g):
单位定义:25℃中,每g组织在反应体系中每分钟催化1 nmol NADH氧化定义为一个酶活力单位。
Rubisco (U/g)=337.6×ΔA测÷W
其中W为样本质量(g),常数337.6由反应体系参数(体积、NADH摩尔消光系数、光径、反应时间等)推导得出,是KTB1480体系专属参数,不适用于其他检测系统。
按蛋白浓度(U/mg prot):
单位定义:25℃中,每mg组织蛋白在反应体系中每分钟催化1 nmol NADH氧化定义为一个酶活力单位。
Rubisco (U/mg prot)=337.6×ΔA测÷Cpr
其中Cpr为蛋白浓度(mg/mL)。蛋白归一化适合在蛋白含量差异较大的样本间进行比较(如不同物种、不同发育阶段),推荐配合BCA蛋白定量试剂盒(亚科因KTD3001)使用。
小结
Rubisco是光合碳固定的唯一入口,也是植物响应逆境、改良光合效率研究的核心靶点。理解它的双重催化特性(羧化/加氧竞争)、了解NADH偶联比色法的信号传递逻辑、清楚方法能测什么和不测什么,是设计一个可靠实验方案的前提。
如果你的下一步是动手操作,可以参考本系列的实验方案篇,了解取样时间控制、UV板选择、ΔA范围判断等实操细节;如果你的课题涉及逆境胁迫或作物光合改良,本系列的热点课题篇提供了更多实验设计参考。
