细菌代谢与菌群研究葡萄糖检测试剂盒使用指南:从抗生素耐药到肠道代谢
葡萄糖检测在微生物和菌群研究中的使用逻辑,和肿瘤细胞实验有一个根本性的差异:细菌对葡萄糖的消耗速率极快,代谢背景更复杂,而且实验目的往往不只是"测消耗量",而是把葡萄糖代谢状态作为切入口,去理解细菌的生存策略或宿主-菌群的代谢互作。
近年来,两个方向的研究让葡萄糖检测在微生物领域的使用变得更加精细:一是细菌自身的碳源代谢与抗生素耐受/耐药机制的关联;二是肠道菌群通过代谢干预宿主糖代谢,进而影响肥胖、糖尿病等代谢性疾病的进程。这篇文章围绕这两个方向,结合具体研究案例,梳理葡萄糖检测在其中的实验角色和操作要点。
细菌碳源代谢与抗生素耐药:一个被低估的联系
氨苄西林操控葡萄糖代谢,切换细菌耐受与耐药状态
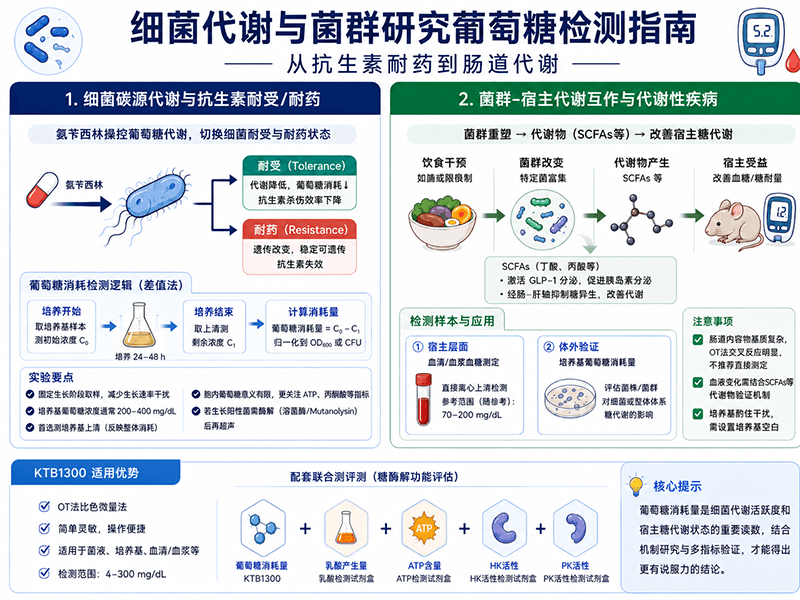
Jiang等人在Science Advances发表的研究提出了一个反直觉的发现:氨苄西林(ampicillin)不只是一种破坏细菌细胞壁的抗生素,它还能主动干预细菌的葡萄糖代谢,而这种代谢干预恰恰是细菌在"耐受(tolerance)"和"耐药(resistance)"两种状态之间切换的关键机制之一。
耐受和耐药是两个容易混淆但机制截然不同的概念。耐药通常涉及遗传突变(如β-内酰胺酶基因的获得),是稳定可遗传的;耐受则是一种非遗传性的表型可塑性,细菌在不改变基因组的前提下进入低代谢、低活性状态,使抗生素的杀伤效率大幅下降——这也是临床上抗生素治疗"明明没有耐药基因、却反复发作"的重要原因之一。
这项研究的核心实验思路之一,就是在不同氨苄西林处理条件下,检测细菌的葡萄糖消耗量变化,以此追踪细菌代谢状态的转变——高葡萄糖消耗对应活跃代谢状态,低消耗则提示细菌进入耐受性低活性状态。葡萄糖消耗量在这里充当的是细菌"代谢活跃度"的间接指示剂,而不只是碳源利用量的简单记录。
这个实验逻辑对后续做抗生素-代谢联合研究的课题组有很强的参考意义:细菌葡萄糖消耗量的下降,不一定意味着生长抑制,也可能意味着耐受状态的形成。仅凭菌落计数判断抗生素效果,会遗漏耐受表型这一层信息。
细菌葡萄糖消耗检测的实验特点
细菌实验和肿瘤细胞实验在葡萄糖检测的操作层面有几处明显差异,值得单独说明。
样本前处理:超声破碎的适用场景与局限
在讨论破碎方案之前,有一个更根本的问题值得先说清楚:测定"细菌胞内游离葡萄糖"在大多数情况下意义有限。细菌摄入葡萄糖后,通过磷酸转糖基转移酶系统(PTS)几乎立即将其磷酸化为6-磷酸葡萄糖,进入糖酵解或合成代谢途径,胞内游离葡萄糖的积累量极低。用比色法试剂盒检测菌体裂解液中的"游离葡萄糖",读数通常接近检测下限,结果噪声大、意义模糊。
因此,细菌实验中葡萄糖检测更有实际意义的终点是培养基中的葡萄糖消耗量:直接离心去除菌体,取培养基上清测定起始与终止浓度之差,反映细菌整体的碳源利用速率。如果课题需要了解胞内代谢状态,更合适的替代指标是ATP含量、丙酮酸浓度或糖原储备,而不是游离葡萄糖。
如果实验确实需要破碎菌体(如测定胞内ATP或其他代谢物并同步测定葡萄糖作为参照),KTB1300的标准破碎方案是收集约500万细菌,冷PBS清洗后加入1 mL PBS,冰浴超声破碎5分钟(功率20%或200 W,超声3秒、间隔7秒,循环30次),12,000 g离心后取上清。
需要注意的是,这套超声参数对革兰氏阴性菌(如大肠杆菌)效果较好,但对革兰氏阳性菌(如金黄色葡萄球菌、肠球菌,以及大多数厚壁菌门肠道菌)效率往往不足——这类细菌的细胞壁含有厚实的肽聚糖层,单纯超声发热风险大且破碎不彻底。专业的微生物代谢实验中,通常在超声前先用**溶菌酶(Lysozyme)或变构酶(Mutanolysin)**进行酶解预处理:溶菌酶切割肽聚糖中的β-1,4-糖苷键,Mutanolysin对厚壁菌门的肽聚糖结构特异性更强。酶解处理后再进行短时超声,破碎效率显著提升,且样本温度更易控制。如果实验涉及多种菌株的横向比较,破碎效率的一致性是保证数据可比性的前提,建议统一验证各菌株在给定方案下的破碎完整性(如通过总蛋白浓度或菌落计数确认破碎率)。
细菌生长速率的干扰
细菌在对数生长期的葡萄糖消耗速率远高于稳定期和衰亡期,因此取样时间点的选择对结果影响极大。同一处理组内如果不同样本处于不同生长阶段,葡萄糖消耗数据的组间变异会很大,甚至掩盖真实的处理效应。标准做法是在实验开始前确认所有样本处于相同的生长阶段(通常以OD₆₀₀校准接种量),并在固定时间点取样。
培养基葡萄糖浓度通常较低
不同于DMEM高糖培养基4500 mg/dL的起始浓度,LB培养基本身不含葡萄糖,M9最小培养基或其他化学限定培养基中外加的葡萄糖浓度通常在0.2%-0.4%(约200-400 mg/dL)。这个浓度范围与KTB1300的检测上限(300 mg/dL)接近,经过一段时间培养后剩余浓度可能已经处于检测范围内,不需要大倍数稀释,但也需要在预实验中确认是否超出线性范围。
肠道菌群研究中的葡萄糖检测
赖氨酸限制饮食通过菌群重塑改善肥胖
Zhao等人在Nature Communications(2025)发表的研究探讨了赖氨酸限制饮食对肥胖的改善效果,核心发现是赖氨酸限制通过富集特定肠道菌群(Parabacteroides goldsteinii)和提升1,4-甲基咪唑乙酸(1,4-methylimidazoleacetic acid)水平,改善了宿主的代谢表型。在评估宿主代谢改善程度时,血糖水平(空腹血糖和葡萄糖耐量)是最直接的代谢功能性读数之一。
这类菌群-代谢互作研究中,葡萄糖检测通常出现在两个层面:一是宿主层面的血清/血浆血糖测定(反映菌群干预对宿主代谢的系统性影响);二是体外验证实验中,测定分离菌株或菌群提取物对细胞葡萄糖消耗的影响(验证菌群代谢物的直接作用)。
理解菌群如何改变宿主血糖,需要在实验设计上意识到这中间存在一个机制桥梁:肠道菌群本身并不直接进入血液,它们调控宿主糖代谢最重要的介质通常是短链脂肪酸(SCFAs),尤其是丁酸(butyrate)和丙酸(propionate)。丁酸是结肠上皮细胞的主要能量来源,同时能激活肠道L细胞分泌GLP-1,间接促进胰岛素分泌;丙酸通过门脉循环进入肝脏,抑制糖异生,降低空腹血糖。菌群在肠道内消耗膳食纤维和碳源产生SCFAs,SCFAs再经肠-肝轴或肠-脑轴调控全身性糖耐量——这才是"菌群干预→血糖变化"这条路径的完整机制链条。
在Zhao等人的研究中,Parabacteroides goldsteinii的富集和1,4-甲基咪唑乙酸水平的提升,背后也涉及类似的代谢物介导机制。这意味着在此类研究里,仅测定宿主血糖往往只能描述表型结果,完整的机制验证还需要配套测定肠道或血清中的SCFAs水平,以及相关代谢物的变化。
KTB1300的血清/血浆检测流程适合第一类场景,样本直接离心澄清后上机,正常啮齿类动物血糖参考值在70-200 mg/dL区间,高脂饮食肥胖模型动物血糖可能显著升高,建议预实验确认是否需要稀释。
肠道葡萄糖:一个特殊的样本类型
如果课题设计涉及肠道内容物或粪便中的葡萄糖水平测定,需要特别注意样本基质的复杂性。肠道内容物中同时含有大量未消化的多糖、寡糖、菌群代谢产生的各类有机酸,以及宿主分泌的消化酶。OT法在这类基质中的特异性问题(总醛糖响应而非单纯葡萄糖)会比在血清样本中严重得多——肠道样本里的其他醛糖来源极其丰富。
如果实验目的是监测肠道葡萄糖吸收后的变化(如肠道葡萄糖转运效率),通常更好的替代方案是测血清/门脉血葡萄糖,而不是直接测肠道内容物。如果确实需要测肠道内容物,应在方法学部分明确说明OT法的这一局限性,并考虑加入HK法平行验证。
微量法在菌群研究中的灵敏度考量
菌群研究中一类常见场景是体外筛选——在不同碳源条件下培养特定菌株,测定其对葡萄糖的偏好性或消耗速率,以此评估菌株的代谢特征。这类实验通常样本量多、单个样本中菌体数量和葡萄糖浓度差异大。
KTB1300葡萄糖检测试剂盒灵敏度4 mg/dL的下限,对于大多数实验室规模的菌株培养实验是够用的,尤其是葡萄糖相对充足的富培养基条件。但在以下情况下需要重新评估:
培养基葡萄糖浓度极低(<10 mg/dL,如模拟肠道低糖环境的限定培养基)或样本前处理引入了大量稀释,实测ΔA值可能低于最低标准品(4 mg/dL)。此时应考虑荧光法葡萄糖检测试剂盒,其检测下限通常可达0.1 µM量级,远低于比色法。
此外,不同菌株的破碎效率差异较大——革兰氏阳性菌细胞壁厚,同样的超声参数下破碎效率可能低于革兰氏阴性菌,导致胞内葡萄糖释放不完全。如果不同菌株之间的比较是实验核心,建议先验证各菌株在给定超声参数下的破碎完整性。
两类研究场景的检测逻辑对比
| 研究方向 | 检测样本 | 测定目标 | 操作要点 |
|---|---|---|---|
| 细菌耐药/耐受机制 | 菌液上清(测培养基消耗)或菌体裂解液(测胞内代谢) | 细菌代谢活跃度、碳源利用状态 | 固定生长阶段取样;以OD₆₀₀归一化;区分胞内/胞外测定目的 |
| 菌群-宿主代谢互作 | 血清/血浆(宿主血糖)或细胞培养基(体外验证) | 菌群干预对宿主糖代谢的系统影响 | 血清直接上机;细胞培养基注意酚红干扰;肠道内容物不推荐直接用OT法 |
细菌和菌群研究给葡萄糖检测带来了与哺乳动物细胞实验不同的挑战——样本类型更多样、代谢背景更复杂、对取样时间点的依赖性更强。在这些场景中,比色法的价值在于它能以较低的操作成本快速获得群体水平的代谢状态读数,适合大批量条件筛选;而需要单菌株精确代谢通量数据的场景,则需要升级到代谢组学或同位素示踪方案。两者并不互斥,在同一个研究里分阶段使用是比较合理的工作流。
