谷胱甘肽过氧化物酶(GSH-Px)是什么?从生物学功能到两种检测原理,一文读懂
适读人群:初次接触GSH-Px检测的研究生、需要了解检测原理以设计实验或撰写方法学的科研人员
核心问题:GSH-Px在生物体内做什么?微量法和DTNB比色法的检测逻辑分别是什么?为什么两种方法能测同一种酶?
读这篇文章之前,先问你一个问题
你在方法学部分写下"检测GSH-Px活性以评估氧化应激水平"——但如果审稿人问你:
- "为什么GSH-Px活性下降代表氧化损伤加剧,而不是氧化清除能力增强?"
- "你用的是NADPH偶联法,但文献里有用DTNB法得到的数据,两者结果可以直接比较吗?"
- "你说NADPH在340 nm的吸光度下降反映GSH-Px活性,但NADPH消耗和GSH-Px之间隔了好几步反应,这条偶联链是怎么保证线性的?"
能答上来吗?
这篇文章的目标,是让你不只会做这个实验,还能在审稿人面前把原理说清楚。
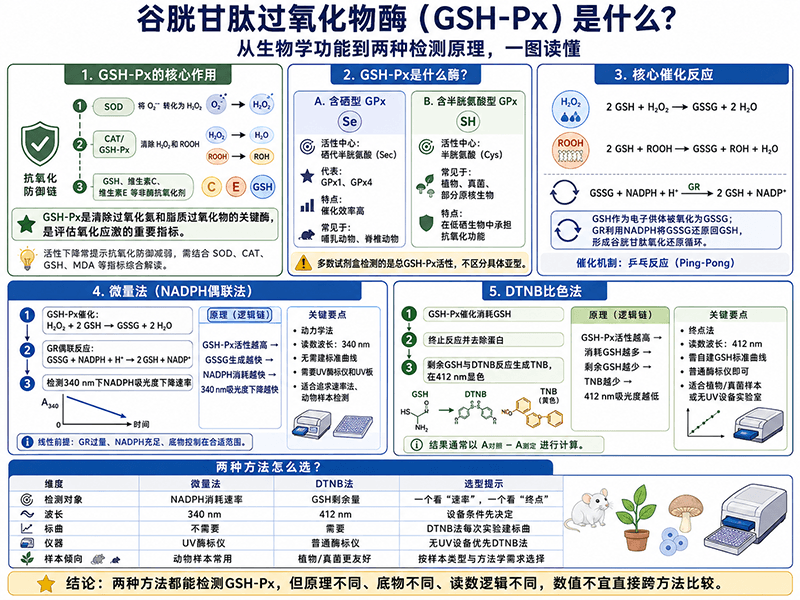
一、GSH-Px是什么?先从氧化应激说起
细胞为什么需要抗氧化系统?
生物体在正常代谢过程中,线粒体电子传递链、细胞色素P450代谢、吞噬细胞的氧化爆发等过程都会持续产生活性氧(Reactive Oxygen Species, ROS),包括超氧阴离子(O₂⁻)、过氧化氢(H₂O₂)、羟自由基(·OH)等。
少量ROS是正常信号分子,参与细胞增殖、免疫防御和基因调控;但过量ROS会攻击DNA、蛋白质和脂质,造成氧化损伤,引发炎症、细胞凋亡,乃至器官功能障碍。
生物体演化出了多层次的抗氧化防御系统来应对这一威胁:
第一层:超氧化物歧化酶(SOD) O₂⁻ + O₂⁻ + 2H⁺ → H₂O₂ + O₂ (把超氧阴离子转化为过氧化氢) 第二层:过氧化氢酶(CAT)和谷胱甘肽过氧化物酶(GSH-Px) H₂O₂ → H₂O + O₂(CAT) H₂O₂ + 2GSH → GSSG + 2H₂O(GSH-Px) (把过氧化氢转化为无害的水) 第三层:非酶类抗氧化剂 维生素C、维生素E、谷胱甘肽(GSH)等
GSH-Px处于这条防线的关键节点——SOD产生的H₂O₂,以及脂质过氧化产生的有机过氧化物(ROOH),最终都要靠GSH-Px和CAT来清除。两者分工略有不同:CAT主要在过氧化物酶体中发挥作用,而GSH-Px广泛分布于细胞质、线粒体等多个亚细胞区室,覆盖范围更广,也是实验中最常检测的清除酶之一。
GSH-Px的两类亚型:含硒的与不含硒的
GSH-Px并不是一个单一的酶,而是一个酶家族(GPx1–GPx8),根据活性中心的不同分为两类:
含硒型(Selenocysteine-containing GPx)
- 活性中心氨基酸:硒代半胱氨酸(Sec,U)
- 代表成员:GPx1(细胞质型,最丰富)、GPx4(磷脂氢过氧化物型)
- 分布:哺乳动物、大多数脊椎动物
- 特点:催化效率极高,对H₂O₂和脂质过氧化物均有活性
- 与硒的关系:硒是此类GSH-Px的必需元素,机体缺硒时GPx1活性显著下降,因此GSH-Px活性可作为评估机体硒营养状态的生化指标
含半胱氨酸型(Cysteine-containing GPx)
- 活性中心氨基酸:半胱氨酸(Cys,C)
- 代表成员:植物GPx、部分真菌GPx
- 分布:植物、真菌、部分原核生物
- 特点:催化效率低于含硒型,但在硒资源匮乏的生物体中承担同等功能
⚠️ 实验设计注意:两款CheKine™ GSH-Px检测试剂盒检测的是总GSH-Px活性,不区分亚型。若研究特定亚型(如GPx4在铁死亡中的作用),需结合亚型特异性抗体或基因敲除验证。
GSH-Px为什么在那么多研究领域都是必测指标?
| 研究领域 | GSH-Px的核心意义 |
|---|---|
| 缺血再灌注损伤 | 再灌注后ROS爆发式产生,GSH-Px活性下降是损伤程度的重要指标 |
| 器官毒理学(肝、肾) | 毒物(酒精、重金属、药物)导致GSH-Px活性降低,氧化损伤加剧 |
| 糖尿病及其并发症 | 高血糖状态下GSH-Px活性下降,参与糖尿病血管损伤的病理机制 |
| 植物抗逆研究 | 干旱、盐碱、重金属胁迫下,植物GSH-Px活性变化反映抗氧化应答强度 |
| 水产养殖与环境胁迫 | 盐度、温度突变时,贝类/鱼类GSH-Px活性是应激评价的核心指标 |
| 硒营养与硒代谢 | GSH-Px是硒的主要生物学载体,其活性直接反映机体有效硒水平 |
| 铁死亡(Ferroptosis) | GPx4是铁死亡的核心调节蛋白,GSH-Px活性是铁死亡研究的必测项目 |
二、GSH-Px催化反应的核心化学
理解检测原理之前,先把GSH-Px本身的催化机制搞清楚。
底物与反应方程式
GSH-Px以**还原型谷胱甘肽(GSH)**为电子供体,催化以下反应:
清除过氧化氢: 2 GSH + H₂O₂ ──GSH-Px──→ GSSG + 2 H₂O 清除有机过氧化物: 2 GSH + ROOH ──GSH-Px──→ GSSG + ROH + H₂O
反应产物**GSSG(氧化型谷胱甘肽)**是有毒的,需要被谷胱甘肽还原酶(GR)在NADPH的驱动下还原回GSH,才能完成"谷胱甘肽氧化还原循环":
GSSG + NADPH + H⁺ ──GR──→ 2 GSH + NADP⁺
这条循环不断消耗NADPH,因此细胞中NADPH的供应(主要来自磷酸戊糖途径)对于维持GSH-Px的持续抗氧化能力至关重要。
催化机制:乒乓反应(Ping-Pong mechanism)
GSH-Px采用乒乓双底物反应机制:
- 氧化半反应:底物(H₂O₂或ROOH)先与活性中心的硒代半胱氨酸(Se-H)反应,将其氧化为亚硒酸形式(Se-OH)
- 还原半反应:GSH依次结合,将Se-OH还原回Se-H,GSH被氧化为GSSG
整个过程中,底物与GSH是依次结合、依次离开,而非同时结合,这也是该酶米氏动力学具有特殊性的原因。
三、微量法(NADPH偶联法)的检测原理
原理概述
微量法不直接检测GSH-Px的底物或产物,而是通过偶联一个辅助反应,将GSH-Px活性转化为可精确测量的NADPH吸光度变化。
完整的偶联反应链如下:
Step 1(目标反应,由GSH-Px催化): H₂O₂ + 2 GSH ──GSH-Px──→ GSSG + 2 H₂O Step 2(偶联反应,由谷胱甘肽还原酶GR催化): GSSG + NADPH + H⁺ ──GR──→ 2 GSH + NADP⁺
关键逻辑:Step 1产生多少GSSG,Step 2就消耗多少NADPH。GSH-Px活性越高→GSSG产生越快→NADPH消耗越快→340 nm处吸光度下降越快。
因此,NADPH在340 nm处的吸光度下降速率(ΔA/min)= GSH-Px催化活性的定量反映。
为什么偶联链能保持线性?
这是理解该方法可靠性的核心问题。前提条件有三:
- GR过量:试剂盒中加入的GR浓度远超催化需求,确保Step 2的速率不成为限速步骤——Step 1产生多快的GSSG,Step 2就能多快地将其转化为NADPH消耗,不存在GSSG积累导致的反馈抑制
- NADPH充足:反应体系中预装足量NADPH,确保在检测时间窗口(10分钟)内,NADPH不会耗尽,始终保持线性消耗
- H₂O₂控制:H₂O₂作为GSH-Px底物,在试剂盒设计浓度下不饱和,确保检测处于酶动力学的线性区间
???? 这也是为什么操作时间窗口严格的原因:GR、NADPH、H₂O₂三者缺一不可地维持着偶联链的线性。样本加入后,必须在规定时间内(10秒和10分10秒)完成两次读数,超时则NADPH消耗进入非线性区,数据不可靠。
信号检测
- 检测波长:340 nm(NADPH的特征吸收峰)
- 信号方向:吸光度随时间下降
- 计算依据:NADPH的摩尔消光系数 ε = 6.22×10³ L/mol/cm(固定物理常数,无需建标准曲线)
- 计算公式(简化版):
GSH-Px (U/g) = 321.6 × (ΔA测定 - ΔA空白) ÷ W
其中W为样本质量(g),321.6为包含反应体积、光径、消光系数和反应时间的综合系数
方法特点总结
| 特点 | 说明 |
|---|---|
| 检测类型 | 动力学法(连续监测吸光度变化速率) |
| 信号物质 | NADPH(间接指示物) |
| 是否需建标准曲线 | 否,基于物理常数直接计算 |
| 操作关键 | 时间窗口严格,加入H₂O₂后必须立即计时 |
| 所需设备 | 能检测340 nm的紫外酶标仪 + 96孔UV板 |
四、DTNB比色法的检测原理
原理概述
DTNB法(5,5'-二硫代双(2-硝基苯甲酸)比色法)采用完全不同的检测逻辑——它不借助偶联反应,而是直接测量GSH-Px催化反应消耗掉多少GSH。
核心反应链如下:
Step 1(酶促反应,由GSH-Px催化,37℃孵育10 min): 有机过氧化物(ROOH)+ 2 GSH ──GSH-Px──→ GSSG + ROH + H₂O (GSH被消耗,体系中剩余GSH减少) Step 2(终止反应): 加入Reagent III(偏磷酸,沉淀蛋白质,终止酶促反应) Step 3(显色反应): 剩余 GSH + DTNB ──→ TNB(黄色,412 nm吸收)+ 混合二硫化物
关键逻辑:GSH-Px活性越高→酶促反应消耗GSH越多→剩余GSH越少→与DTNB反应产生的TNB越少→412 nm吸光度越低。
因此,测定孔与对照孔之间412 nm吸光度的差值(ΔA测定 = A对照 - A测定),正比于被GSH-Px消耗的GSH量,也就是GSH-Px的催化活性。
???? 注意信号方向:DTNB法中信号越强(吸光度越高)代表剩余GSH越多,也就是GSH-Px活性越低;信号越弱代表GSH被消耗越多,GSH-Px活性越高。这与微量法(吸光度下降代表酶活高)的信号方向逻辑一致,但物理含义相反,计算时要用A对照减去A测定,而不是直接用A测定的值。
为什么要设"对照管"?
DTNB法的实验设计中有一个容易被忽视的细节:除了测定管,还有一个对照管——对照管中,先加入Working Reagent(GR底物体系)、孵育,再加入终止剂(Reagent III)之后才加入样本。
这样设计的原因是:对照管中GSH-Px来不及催化反应(因为终止剂已经先行沉淀蛋白),对照管代表的是"完全没有GSH-Px消耗时体系中GSH的总量"。
用对照管吸光度减去测定管吸光度,得到的差值正好代表"GSH-Px在10分钟酶促反应中消耗掉的GSH量",排除了样本内源物质对DTNB显色的背景干扰。
标准曲线的作用
由于不同实验室的仪器响应、试剂批次略有差异,DTNB法要求每次实验自建标准曲线:用已知浓度的GSH标准品(6.25–400 nmol/mL,7个梯度)与DTNB反应,以ΔA为横轴、标准品浓度为纵轴作图,得到线性方程后代入样本ΔA换算实际GSH消耗量,再根据反应体积换算为酶活。
信号检测
- 检测波长:412 nm(TNB的特征吸收峰,可见光范围,黄色)
- 信号方向:测定孔吸光度低于对照孔(活性越高差值越大)
- 计算依据:标准曲线方程(每次实验自建,典型R²≥0.999)
- 所需设备:普通酶标仪 + 普通透明96孔板
方法特点总结
| 特点 | 说明 |
|---|---|
| 检测类型 | 终点法(固定时间点读取最终吸光度) |
| 信号物质 | TNB(GSH与DTNB的反应产物,直接指示物) |
| 是否需建标准曲线 | 是,每次实验必须自建 |
| 操作关键 | 加样顺序、孵育时间和终止时机必须严格统一 |
| 所需设备 | 普通酶标仪 + 普通酶标板 |
五、两种方法的原理对比与选择逻辑
原理层面的本质差异
| 维度 | 微量法(NADPH偶联法) | DTNB比色法 |
|---|---|---|
| 检测对象 | NADPH消耗速率(间接) | GSH剩余量(直接) |
| 反应类型 | 动力学(速率法) | 终点法 |
| 信号物质 | NADPH(340 nm,紫外) | TNB(412 nm,可见光) |
| 偶联步骤 | 需要(引入GR和NADPH) | 不需要(直接显色) |
| 底物 | H₂O₂ | 有机过氧化物(叔丁基过氧化氢等) |
| 活性单位定义 | 每分钟催化1 nmol NADPH氧化 | 每分钟催化1 nmol GSH氧化 |
为什么两种方法的数值不能跨方法比较?
这个问题本质上有两个层面:
层面一:活性单位的定义不同,计量推导路径也不同
两种方法检测的"信号分子"不同,背后的化学计量推导逻辑各自独立:
微量法(NADPH偶联法):GSH-Px催化1 mol H₂O₂消耗2 mol GSH,产生1 mol GSSG;GR再催化1 mol GSSG消耗1 mol NADPH还原回2 mol GSH。因此,ΔNADPH : ΔGSSG = 1:1,ΔNADPH : ΔGSHconsumed = 1:2。活性单位定义为"每分钟催化1 nmol NADPH氧化"。
DTNB比色法:GSH-Px催化有机过氧化物消耗GSH,剩余GSH与DTNB发生显色反应,计量关系为1 mol GSH → 1 mol TNB(1:1)。标准曲线以已知GSH浓度定标,ΔA直接换算为"消耗的GSH量(nmol/mL)"。活性单位定义为"每分钟催化1 nmol GSH氧化"。
两种方法的活性单位在化学上虽然都能追溯到同一个GSH-Px催化事件,但检测信号分子不同(NADPH vs TNB)、计量推导路径不同、底物也不同(H₂O₂ vs 有机过氧化物),因此数值之间不存在简单的换算系数,不能跨方法直接比较。
层面二:底物不同,检测的GSH-Px活性谱不同微量法用H₂O₂作为底物,主要反映针对H₂O₂的GSH-Px活性;DTNB法用有机过氧化物作为底物,反映的是针对脂质过氧化物的活性。两者在大多数生理情境下高度相关,但在特定病理状态(如GPx4特异性失活、脂质过氧化物积累)中可能出现分离。
实际结论:同一研究中始终使用同一方法,方法学部分写清楚货号和检测原理,与文献比较时说明所用方法即可,无需换算。
六、两种检测原理的常见疑问解答
Q:微量法测的是NADPH消耗,那体系中是否有内源性NADPH酶会干扰结果?
A:确实存在这个风险。样本中可能含有能消耗NADPH的其他酶,如内源性谷胱甘肽还原酶(GR)、硫氧还蛋白还原酶(TrxR)、醛脱氢酶等,它们都会造成340 nm吸光度非特异性下降。
商用试剂盒的处理方式:设置"试剂空白管"——用等体积去离子水代替样本,检测同样Working Reagent体系下NADPH的自然衰减速率,用ΔA测定减去ΔA空白进行背景校正。这个方式操作简便,适用于大多数样本。
高背景样本的注意事项:对于大鼠肝脏、心肌等代谢活跃、内源酶丰富的组织,若遇到空白管信号异常偏高(ΔA空白接近甚至超过ΔA测定),说明样本基质中存在显著的非GSH-Px来源的NADPH消耗,此时可尝试:
- 适当稀释样本,降低背景酶的浓度干扰;
- 设置"样本空白管"(加样本但不加H₂O₂),单独评估样本基质本身的NADPH消耗背景;
- 与说明书中的正常ΔA范围对照,若严重偏离,建议联系技术支持(400-6800-830)排查。
因此,空白管不能省略,且遇到异常数据时应首先排查空白管信号是否正常。
Q:DTNB法中,为什么要在离心取上清后再转移到96孔板显色,而不是直接在EP管里读数?
A:Reagent III(偏磷酸)加入后会沉淀大量蛋白质,导致溶液浑浊,直接在EP管或96孔板检测会产生严重的散射光干扰。离心(5000 g,4℃,10 min)后取上清,目的正是去除蛋白沉淀,获得清澈的溶液,保证吸光度读数准确。
Q:两种方法对GSH-Px的全部8个亚型(GPx1–GPx8)都能检测吗?
A:理论上,两种方法检测的是具有催化活性的总GSH-Px,主要反映样本中GPx1(细胞质型,含量最丰富)的活性。GPx4(磷脂氢过氧化物型)、GPx3(分泌型)等其他亚型也有贡献,但丰度相对低。如需区分特定亚型的活性,目前尚无商品化的亚型特异性活性检测方案,通常需要结合基因敲除、特异性抑制剂(如RSL3靶向GPx4)来间接判断。
Q:GSH-Px活性降低一定意味着氧化损伤加重吗?
A:不完全是,需要结合背景解读:
- 如果GSH-Px活性降低同时伴随MDA(丙二醛)升高、GSH含量下降,则提示氧化损伤加重;
- 如果GSH-Px活性轻度降低但其他指标正常,也可能是底物(GSH、硒)供应不足导致的酶活力下降,而非损伤;
- 铁死亡情境下GPx4活性的特异性丧失与总GSH-Px活性变化可能不完全一致。
建议:GSH-Px活性应与SOD、CAT、MDA、GSH等指标联合解读,不建议单独报告GSH-Px活性作为氧化应激的唯一依据。
Q:样本采集和前处理阶段有哪些容易忽视的干扰因素?
A:样本处理是GSH-Px检测误差最集中的环节,以下几类问题在实际实验中高频出现:
① 组织/细胞裂解方式如第一篇选购指南中所述,RIPA、NP40等含SDS或去氧胆酸钠的裂解液会破坏GSH-Px的天然四聚体构象,导致活性丧失。应严格使用试剂盒配套的Assay Buffer(微量法)或Extraction Buffer(DTNB法)进行匀浆或超声裂解。超声参数(200 W,3 s开/7 s停)也不建议随意调整,功率过高同样会导致酶失活。
② 样本中的氧化剂残留组织在采集过程中如接触到H₂O₂、次氯酸钠(消毒剂)等氧化性物质,会直接消耗样本中的GSH底物,造成后续检测中GSH-Px"表观活性"偏高的假象(GSH被预消耗,与DTNB反应的信号降低)。建议样本采集全程避免氧化性试剂污染,用预冷PBS彻底清洗组织后再匀浆。
③ 血清/血浆样本的采集管选择血清样本可直接检测,通常是最稳定的液体样本类型。若使用血浆,建议在说明书推荐范围内选择抗凝管,并记录抗凝剂种类写入方法学——不同抗凝剂对酶活的潜在影响在文献中存在讨论,若遇到血浆与血清检测结果差异较大的情况,建议平行比较两种样本类型以排除采集方式干扰。
④ 样本保存时间与冻融次数GSH-Px对反复冻融较为敏感,建议新鲜样本当天检测;如需保存,-80℃单次冻融,避免分装不足导致多次开管。组织匀浆液在冰上放置超过2–3小时后,酶活会出现不同程度的自然衰减,应尽量缩短样本制备到上机检测的时间间隔。
七、快速选型总结
搞清楚原理之后,选择哪款试剂盒的逻辑就很清晰了:
- 需要动力学速率法、无需建标曲、样本为动物来源 → 选 KTB1640(微量法)
- 需要终点法、有植物/真菌样本、实验室无UV酶标仪 → 选 KTB1641(DTNB比色法)
- 不确定哪款适合自己的实验 → 参考《GSH-Px活性检测试剂盒怎么选?微量法 vs DTNB比色法,避开这6个坑再下单》的完整选购指南
本产品仅供科学研究使用,不适用于临床诊断。
