从肿瘤氧化应激到病毒代谢劫持:为什么NADP⁺/NADPH正在成为这些课题的核心检测指标?
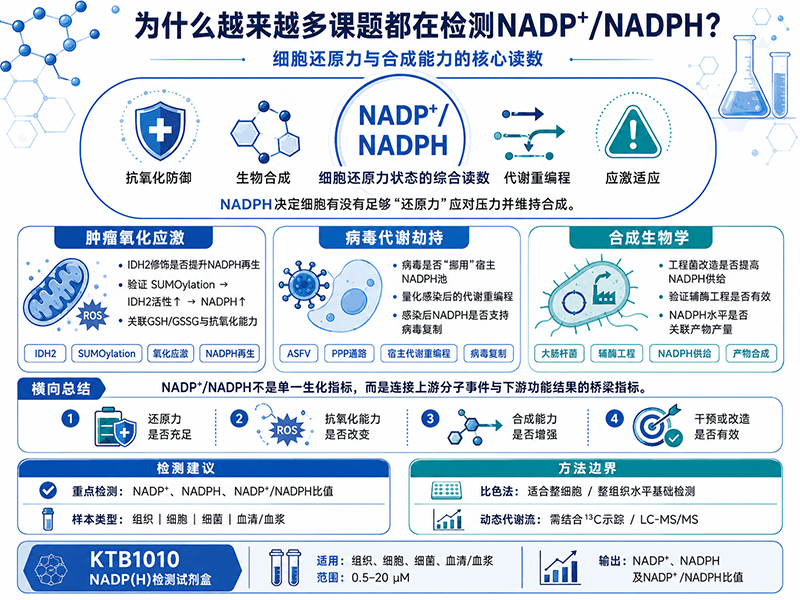
如果说NAD(H)是细胞的"能量账本",NADP(H)更像是细胞的"应急储备金"——它的核心功能不是产能,而是在氧化应激来袭时提供还原力,同时为脂质、核酸等生物合成反应持续供给原料。正因如此,NADP⁺/NADPH的失衡出现在一系列看似不相关的研究领域里:肿瘤细胞的氧化应激适应、病毒感染对宿主代谢的系统性重编程、以及工业微生物的辅酶工程改造。
这篇文章从三个有代表性的课题方向切入,说清楚每个方向为什么要测NADP⁺/NADPH、检测结果在机制论证中承担什么角色。
方向一:IDH2的SUMOylation修饰——氧化应激下NADPH再生的调控节点
课题背景
异柠檬酸脱氢酶2(IDH2)是线粒体内NADPH再生的核心酶之一,催化异柠檬酸氧化脱羧生成α-酮戊二酸,同时将NADP⁺还原为NADPH。线粒体NADPH是谷胱甘肽还原酶的驱动力,维持线粒体内GSH/GSSG比值,直接决定线粒体抵抗氧化损伤的能力。
一项发表于Biochemical and Biophysical Research Communications的研究(Kim等,2020)发现,氧化应激条件下IDH2会发生SUMO化修饰(SUMOylation)——这是一种类泛素化的蛋白质翻译后修饰,通过将SUMO蛋白共价连接到靶蛋白的特定赖氨酸残基上来调控靶蛋白功能。研究表明,IDH2的SUMOylation在氧化应激下显著增强,而这种修饰正向调控了IDH2的酶活性,使细胞在应对氧化压力时能够加速NADPH的再生。当SUMOylation位点突变、修饰被阻断后,IDH2活性受损,NADPH水平下降,细胞对氧化应激的耐受能力随之减弱。
这个发现把翻译后修饰和辅酶代谢连接了起来:细胞感知到氧化应激信号后,通过增强IDH2的SUMOylation来上调NADPH供给,这是一条此前未被充分认识的氧化应激适应机制。
为什么这个方向需要检测NADP⁺/NADPH?
这个课题的核心论点是:SUMOylation增强IDH2活性,进而提升细胞内NADPH水平。"进而"这两个字在论证上需要实验数据支撑——如果只有IDH2酶活性的体外测定,证明的是修饰影响了酶的催化效率,但不能证明这个效率变化在细胞内转化成了可检测的NADPH含量变化。
需要的检测逻辑是:
- 正向验证:在氧化应激条件下,SUMOylation增强的细胞中NADP⁺/NADPH比值是否向NADPH一侧偏移(即NADPH相对升高,NADP⁺相对降低)
- 反向验证:SUMOylation位点突变后,NADPH水平是否下降,NADP⁺/NADPH比值是否升高
- 功能关联:NADPH水平变化是否与GSH/GSSG比值、细胞存活率在应激条件下的变化存在相关性
这三个层面的数据构成一条完整的因果链:SUMOylation→IDH2活性↑→NADPH↑→氧化应激耐受↑。缺少NADPH的直接定量数据,这条链条在中间断掉了。
NADP⁺/NADPH的比值在这里不只是一个辅助指标,而是连接分子修饰和细胞表型的必要中间节点。
从实验角度,这类课题的样本通常是培养细胞(过表达或敲减SUMO修饰相关酶的细胞系,如SENP2敲减细胞),在H₂O₂或其他氧化应激处理前后收集,提取NADP⁺和NADPH,按细胞数归一化,报告比值变化。KTB1010的检测范围(0.5–20 µM)和细胞样本的NADPH浓度范围通常匹配,起始量1×10⁶个细胞即可。
研究启发
IDH2-SUMOylation这个课题框架,把翻译后修饰研究和代谢组学指标连接了起来,开辟了一条用辅酶水平验证修饰功能的实验思路。类似的逻辑可以延伸到:其他NADPH再生酶的翻译后修饰研究(如G6PD的乙酰化、磷酸化对PPP通量的调控);肿瘤细胞在放化疗诱导的氧化应激下NADPH再生能力的动态变化;以及靶向IDH2突变体的抗肿瘤药物(如AG-221/Enasidenib)对细胞NADPH代谢的影响评估。这些方向都需要NADP⁺/NADPH的直接定量来桥接分子机制和细胞功能。
方向二:非洲猪瘟病毒劫持宿主嘧啶代谢——病毒如何"挪用"NADPH
课题背景
非洲猪瘟(ASF)是全球养猪业面临的毁灭性疫病,非洲猪瘟病毒(ASFV)至今没有有效疫苗,防控依赖扑杀。一项2025年发表于Journal of Virology的研究(Zheng等,Song等,2025)揭示了ASFV感染宿主细胞后如何系统性重编程宿主代谢来服务病毒自身的复制需求。
研究发现,ASFV对宿主细胞代谢进行了重编程,以产生病毒复制所必需的基础物质。具体来说,ASFV操控宿主核苷酸生物合成通路,一方面为DNA合成提供底物,另一方面获取应对氧化应激所需的还原力。嘧啶合成的从头合成通路(de novo synthesis)高度依赖谷氨酰胺和天冬氨酸作为氮源,同时需要NADPH为二氢乳清酸脱氢酶之外的还原步骤提供电子。ASFV通过上调宿主PPP通路活性来扩大NADPH供给,把原本属于宿主细胞用于抗氧化防御的NADPH池,转为服务病毒DNA复制的核苷酸合成。
这是一个典型的"代谢劫持"模式:病毒不直接合成NADPH,而是通过操控宿主的代谢网络,让宿主细胞为病毒生产所需的还原力。阐明ASFV如何劫持感染细胞内的核苷酸生物合成,为对抗非洲猪瘟的创新策略铺平了道路。
为什么这个方向需要检测NADP⁺/NADPH?
这个课题的核心问题是:ASFV感染后,宿主细胞的NADPH代谢发生了什么变化?检测NADP⁺/NADPH在这里承担的是代谢重编程的直接表型读出。
论证ASFV"劫持"了NADPH池,需要的数据层次包括:
- 感染前后的NADP⁺/NADPH比值变化:ASFV感染细胞后,如果PPP通路上调,NADPH供给增加,应该能观察到NADP⁺/NADPH比值向NADPH一侧偏移
- 病毒复制量与NADPH水平的相关性:NADPH供给充足时病毒滴度是否更高;用PPP抑制剂(如6-AN)阻断NADPH再生后,病毒复制效率是否下降
- 抗氧化系统的竞争性消耗:感染细胞中GSH水平是否下降,提示NADPH被优先分配给核苷酸合成而非谷胱甘肽还原
**NADPH不够用,宿主抗氧化防线就会崩溃;NADPH被病毒"挪走",病毒核酸合成就有了充足的还原力。**这个竞争关系的直接证据,需要NADPH的定量数据来支撑,不是PPP通路酶表达的间接推断可以替代的。
从实验角度,ASFV研究通常在猪肺泡巨噬细胞(PAM)或猪骨髓细胞(pBMDM)等原代细胞,以及Vero细胞等允许ASFV复制的细胞系中进行。这些细胞样本按1×10⁶个细胞起始提取,完全在KTB1010的适用范围内。需要注意的是,ASFV是高度危险的动物病原,相关实验必须在符合生物安全要求的设施内进行。
研究启发
病毒代谢劫持是近年来病毒学研究的热点方向,不限于ASFV。新冠病毒(SARS-CoV-2)、流感病毒、疱疹病毒等均被发现能不同程度地重编程宿主细胞的碳代谢和氧化还原代谢。NADP⁺/NADPH检测作为宿主氧化还原状态的综合读数,在病毒-宿主代谢互作研究中具有普适性的应用价值——无论具体病毒如何操控PPP通路或NADPH消耗途径,比值的变化方向都能提供宿主代谢响应的第一手信息。此外,以代谢通路为靶点的抗病毒策略(如PPP抑制、谷氨酰胺拮抗)的效果评估,也需要NADPH水平的定量数据作为药效指标。
方向三:大肠杆菌辅酶工程——NADPH定量是合成生物学的效率刻度盘
课题背景
1,4-丁二胺(腐胺,putrescine)是重要的工业原料,用于聚合物、医药、农化品和表面活性剂的生产。与化学合成相比,微生物发酵生产1,4-丁二胺具有更好的绿色经济性,但生产效率一直是瓶颈。
一项发表于Molecules的研究(Li等,2024)系统性地解析了辅酶PLP(磷酸吡哆醛)和NADPH对大肠杆菌合成1,4-丁二胺的限速作用。合成1 mol 1,4-丁二胺需要消耗2 mol NADPH,因此在大肠杆菌高效合成1,4-丁二胺时,必须考虑NADPH的平衡问题。研究通过优化PLP和NADPH合成相关基因的表达,使胞内PLP和NADPH水平同步提升,最终重组菌株NAP19以葡萄糖为碳源,1,4-丁二胺的产量达到272 mg/L·DCW,比出发菌株提高了79%。
这个研究的核心逻辑是:目标产物的合成效率受限于辅酶供给,而辅酶供给可以通过基因工程手段主动调控。NADPH不再只是一个代谢副产物,而是一个可以被精确设计的"生产要素"。
为什么这个方向需要检测NADP⁺/NADPH?
合成生物学课题中,检测NADP⁺/NADPH的核心目的和疾病研究有所不同——这里不是为了证明"哪里出了问题",而是为了量化工程改造是否真正提升了辅酶供给,以及辅酶水平的提升和产物产量之间的量效关系。
具体需要的数据包括:
- 工程菌株vs出发菌株的NADPH水平对比:基因优化后NADPH是否确实在细胞内积累,排除转录水平变化但代谢通量未实际改变的情况
- NADPH水平与产物产量的相关性:不同基因组合的菌株,NADPH水平和1,4-丁二胺产量是否呈正相关,为后续菌株筛选提供代谢指标
- 发酵过程中的动态监测:在不同发酵时间点取样,追踪NADPH水平的动态变化,找到NADPH供需平衡的关键时间窗口
对合成生物学课题来说,NADPH含量是评估代谢工程改造效果最直接的胞内辅酶指标,测不准这个数,工程改造的逻辑链条就缺了一个关键的验证节点。
细菌样本的提取方式和细胞样本类似,KTB1010同样适用:收集约1×10⁶个菌,冷PBS洗涤,超声破碎(冰浴全程),按NADP提取液和NADPH提取液分别提取,4℃高速离心后取上清检测。
研究启发
辅酶工程是合成生物学中提升目标代谢物产量的通用策略,不限于1,4-丁二胺这一个产品。凡是合成路径中有NADPH消耗步骤的产物——包括氨基酸(谷氨酸、脯氨酸)、有机酸、萜类化合物、生物碱——都面临类似的NADPH供需平衡问题,都需要在菌株改造前后定量检测胞内NADPH水平来验证工程效果。 此外,在代谢流分析中,NADP⁺/NADPH比值是判断PPP通路活性(NADPH主要来源)和TCA循环中IDH1/IDH2活性的间接指标,与13C同位素示踪数据结合,能对菌株的代谢网络给出更完整的描述。
横向视角:三个方向共同指向的检测逻辑
把这三个课题放在一起看,NADP⁺/NADPH检测在不同场景里承担的角色各有侧重,但底层逻辑一致:
| 课题方向 | 检测NADP⁺/NADPH的核心目的 |
|---|---|
| IDH2/SUMOylation(肿瘤氧化应激) | 验证翻译后修饰→酶活性→辅酶水平这条因果链的中间节点 |
| ASFV代谢劫持(病毒学) | 量化病毒感染对宿主NADPH池的重编程程度,作为代谢劫持的直接表型证据 |
| 大肠杆菌辅酶工程(合成生物学) | 确认基因工程改造切实提升了胞内NADPH供给,建立辅酶水平与产物产量的量效关系 |
三个方向的共同点是:NADP⁺/NADPH的直接定量,在论证结构中处于上游分子事件和下游功能结果之间的桥接位置。上游的基因/蛋白表达数据能说明"合成或消耗能力变了",下游的GSH总量、脂质合成速率等功能指标能说明"氧化防御或合成能力变了",而NADPH的直接含量测定能说明"辅酶水平本身变了"——三个层次在论证链条上各有位置,互为补充,不能互相替代。
需要说明的是,比色法测定的是某一时刻细胞内NADPH的静态浓度(pool size),而非NADPH的动态生成速率(flux)。这两个概念在实验解读上有根本区别:静态浓度不变,不代表代谢通量没有改变——有可能是PPP上调加速了NADPH生成,同时核苷酸合成等下游途径同步消耗了等量NADPH,净浓度看起来持平,但代谢重编程已经发生。如果课题需要回答"NADPH的生成速率是否改变了",需要配合¹³C同位素示踪的代谢流分析(MFA),比色法无法提供这个维度的数据。
值得一提的是,NADPH研究还有几个体量更大的应用领域没有在本文展开:肿瘤的铁死亡(ferroptosis)——GPX4的活性高度依赖GSH,而GSH的再生需要NADPH,铁死亡研究中NADPH水平是评估细胞氧化还原状态的核心指标之一;脂肪肝与代谢综合征——肝脏脂肪酸从头合成的大量NADPH消耗,使NADP⁺/NADPH失衡成为非酒精性脂肪性肝病(NAFLD)的代谢特征之一;植物逆境应答——干旱、高温、重金属胁迫下,植物细胞NADPH供给能力的维持与否直接关系到活性氧的清除效率。这些方向都涉及组织或细胞样本的NADP⁺/NADPH定量,比色法是其中最便捷的检测手段之一。
小结
从IDH2的翻译后修饰调控,到ASFV对宿主还原力的系统性劫持,再到工业微生物的辅酶供给工程,NADPH始终处于这些故事的核心位置——它是细胞在逆境中维持还原力的最后防线,是病毒复制所需核苷酸合成的电子供体,也是微生物高效产出目标化合物的能量货币。
检测NADP⁺/NADPH,是把这些机制从假说落到实验数据的必要一步。比色法在这个过程中的定位是:以最低的设备门槛给出整细胞水平的辅酶总量和比值,适合大批量样本的表型筛查、干预效果评估和初步机制验证。对于投向高分期刊的代谢机制研究,审稿人通常会期望看到液相色谱-串联质谱(LC-MS/MS)的数据作为定量验证或补充——LC-MS/MS能直接通过质荷比区分结构高度相似的辅酶(如NADH与NADPH),灵敏度更高,也不依赖酶的底物特异性。比色法和LC-MS/MS不是替代关系,而是适用于不同研究阶段和期刊要求的互补工具:前者效率高、门槛低,适合建立基础表型;后者精度高、信息量大,适合机制深挖和顶刊投稿。
CheKine™ KTB1010(辅酶Ⅱ NADP(H)含量检测试剂盒)支持动植物组织、细胞、细菌和血清(浆)四类样本,检测范围0.5–20 µM,可同时定量NADP⁺和NADPH并计算比值,适合上述各课题方向的整细胞水平基础检测需求。
参考文献
Kim H, et al. SUMOylation enhances the activity of IDH2 under oxidative stress. Biochemical and Biophysical Research Communications, 2020, 533(4): 851–857. https://doi.org/10.1016/j.bbrc.2020.09.082
Zheng Z, Song et al. African swine fever virus hijacks host pyrimidine metabolism to promote viral replication. Journal of Virology, 2025, 99(11): e00985-25. https://doi.org/10.1128/jvi.00985-25
Li M, et al. Increasing 1,4-Diaminobutane Production in Escherichia coli by Optimization of Cofactor PLP and NADPH Synthesis. Molecules, 2024, 29(13): 3094. https://doi.org/10.3390/molecules29133094
