从慢性肾病到肿瘤代谢:为什么越来越多课题把NAD⁺/NADH列为必测指标?
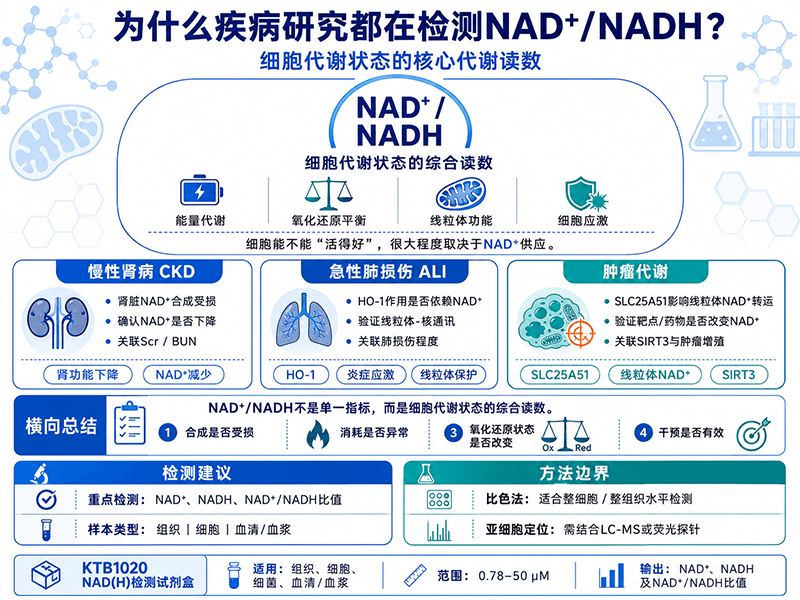
如果你最近在读氧化还原代谢相关的文献,应该不难发现一个趋势:越来越多不同疾病方向的课题开始把NAD⁺/NADH列为核心检测指标。慢性肾病、脓毒症相关急性肺损伤、肿瘤代谢——这些领域看起来相距甚远,但在NAD⁺这个节点上有一个共同的逻辑:细胞能不能活、活得好不好,在很大程度上取决于NAD⁺的供应水平。
这篇文章梳理三个有代表性的课题方向,说清楚每个方向为什么要测NAD⁺/NADH、测出来能回答什么问题。不是文献综述,而是从实验设计的角度解释检测的必要性。
方向一:慢性肾病中的NAD⁺合成障碍
课题背景
慢性肾病(CKD)是全球性公共卫生问题,但它的代谢机制长期以来研究得不够深入。一项发表于Frontiers in Physiology的研究(Li等,2021)系统性地检测了CKD大鼠模型中肾脏NAD⁺和NADH的含量,以及NAD⁺三条生物合成通路中关键酶的表达变化,提供了一个把NAD⁺代谢障碍和CKD病理进程联系起来的实验框架。
研究者建立了两种CKD大鼠模型(5/6肾切除模型和腺嘌呤饲喂模型),通过血清肌酐(Scr)和尿素氮(BUN)确认肾功能损害,同时用NGAL升高、肾小球肥大和肾纤维化标志物确认病理改变。在这个基础上检测了两种模型肾脏中的NAD⁺和NADH含量,结果发现两者在CKD模型中同步下降,而且NAD⁺水平与Scr和BUN呈负相关——肾功能越差,NAD⁺越低。
进一步的机制分析聚焦在NAD⁺的三条合成通路上:从头合成通路(de novo pathway)、补救合成通路(salvage pathway)和Preiss-Handler通路。研究发现,从头合成通路的关键酶QPRT,以及补救通路的NMNAT1和NMNAT3,在两种CKD模型中均显著下调。NAD⁺消耗端的去乙酰化酶SIRT3和CD38也同步下降,提示整个NAD⁺代谢网络在CKD状态下发生了系统性紊乱,而不只是某一个通路出了问题。
为什么这个方向需要检测NAD⁺/NADH?
这个课题的核心问题是:CKD状态下肾脏的能量代谢到底出了什么问题?NAD⁺/NADH的绝对含量和比值是回答这个问题最直接的表型数据。
光测NAD⁺合成酶的表达,只能说明"合成能力下降了",不能证明"NAD⁺真的减少了"。 酶的表达变化不一定和NAD⁺含量变化等比例对应——有可能合成减少了,但消耗也减少了,总量变化不显著。直接检测NAD⁺和NADH的含量,才能给出肾组织实际的辅酶水平,和功能指标(Scr、BUN)之间的相关性分析也才有数据基础。
从实验设计的角度,这类课题通常需要:肾组织匀浆样本(20 mg起始量),分别提取NAD⁺和NADH,报告绝对含量和NAD⁺/NADH比值,按组织质量(nmol/g)归一化,同时和肾功能指标做相关性分析。
研究启发
这个课题框架对以下研究方向有参考价值:其他慢性代谢性疾病的器官NAD⁺代谢研究(糖尿病肾病、肝纤维化、心肌病等);NAD⁺前体补充(NMN、NR)干预慢性疾病的机制研究;以QPRT或NMNAT家族为靶点的药物研发。这些方向都需要在干预前后对比器官组织中的NAD⁺和NADH水平,检测NAD⁺/NADH是评估干预效果的必要指标。
方向二:HO-1通过NAD⁺调控线粒体-核通讯抵抗急性肺损伤
课题背景
脓毒症诱发的急性肺损伤(ALI)是重症监护领域死亡率极高的临床问题,目前没有特异性的治疗手段。血红素加氧酶-1(HO-1)是一个已知的内源性保护蛋白,但它究竟通过什么机制发挥肺保护作用,一直没有完整的答案。
一项发表于Inflammation Research的研究(Yu等,2022)把HO-1的保护作用和NAD⁺水平联系了起来。研究者在内毒素诱导的ALI模型中发现,HO-1通过调控NAD⁺水平来维持线粒体-核通讯(mitonuclear communication)的正常运行,而这条通讯链路的核心中介正是NAD⁺——线粒体把细胞代谢状态通过NAD⁺/NADH比值传递给核基因组,驱动适应性基因表达。
具体来说,研究发现HO-1激活后NAD⁺水平上升,线粒体蛋白失衡(mitonuclear protein imbalance)和线粒体未折叠蛋白反应(UPRmt)得到改善,肺损伤减轻。当PGC1α/PPARγ信号通路被抑制时,NAD⁺水平下降,线粒体-核通讯紊乱加重,肺损伤恶化。这条HO-1→NAD⁺→PGC1α/PPARγ→mitonuclear communication的链路,把一个已知的保护蛋白和NAD⁺代谢、线粒体功能连接成了一个完整的机制通路。
为什么这个方向需要检测NAD⁺/NADH?
这个课题的核心逻辑是:HO-1的保护效果"依赖于"NAD⁺水平。要证明这个依赖关系,需要在以下几个层面上都有NAD⁺的数据支撑:
第一,HO-1激活/抑制前后,肺组织NAD⁺水平是否确实发生了对应变化(上调HO-1→NAD⁺升高;抑制PGC1α/PPARγ→NAD⁺下降);
第二,NAD⁺的变化幅度和肺损伤程度之间是否存在量效关系;
第三,外源补充NAD⁺前体是否能绕过HO-1,直接改善线粒体-核通讯,这是验证NAD⁺为核心中介的经典实验设计。
这三个层面的数据,都需要用能同时检测NAD⁺和NADH、给出比值的方法来支撑。仅有基因或蛋白表达层面的数据是不够的——HO-1上调了NAMPT(NAD⁺合成酶),这只能说明合成能力可能增强;直接测NAD⁺含量才能证明结果。
从实验角度,这类课题的样本通常来自肺组织(大鼠或小鼠ALI模型),以及LPS处理的肺泡上皮细胞系(如A549)。两种样本类型都在KTB1020的适用范围内,组织按20 mg起始,细胞按1×10⁶起始。
研究启发
HO-1和NAD⁺的联系,在脓毒症相关脏器损伤研究里打开了一个新的机制维度。肝脏、肾脏、心脏的脓毒症损伤中,类似的线粒体-核通讯紊乱机制是否也存在?HO-1-NAD⁺轴是否在不同器官之间具有普适性?这些问题都是可以顺着这篇文章的框架延伸出来的课题。更广泛地说,任何涉及线粒体功能障碍的炎症或应激模型,NAD⁺/NADH比值都是描述线粒体能量代谢状态的基础指标,值得纳入检测体系。
方向三:SLC25A51与线粒体NAD⁺转运——肿瘤代谢的新靶点
课题背景
线粒体是细胞内NAD⁺消耗最集中的场所,但哺乳动物细胞如何把胞质中的NAD⁺转运进线粒体,长期是个悬而未决的问题。直到SLC25A51被鉴定为哺乳动物细胞的线粒体NAD⁺转运体,这个问题才有了答案。
一项发表于Cell Death & Differentiation的研究(Luo等,2023)把SLC25A51的功能研究推进到了肿瘤背景下。研究发现SLC25A51在多种肿瘤中表达上调,敲减SLC25A51会导致线粒体NAD⁺输入减少,进而使SIRT3(线粒体去乙酰化酶)功能受损,线粒体蛋白乙酰化水平异常升高。乙酰化升高进一步影响了脯氨酸合成关键酶P5CS的活性,导致脯氨酸含量下降,最终抑制肿瘤细胞增殖。
这个研究还发现FDA批准的药物氟达拉滨磷酸盐(fludarabine phosphate)能够结合并抑制SLC25A51功能,与阿司匹林联用时具有协同抗肿瘤效果。SLC25A51从一个基础生物学问题(NAD⁺怎么进线粒体)变成了一个具有药物开发前景的癌症靶点。
为什么这个方向需要检测NAD⁺/NADH?
这个课题的核心论点是:SLC25A51的功能状态直接决定线粒体NAD⁺的可用量,进而影响SIRT3活性和下游的蛋白乙酰化修饰。检测NAD⁺/NADH在这里承担的是因果链验证的功能。
具体来说需要的数据包括:SLC25A51敲减后线粒体NAD⁺含量是否确实下降(而非胞质NAD⁺下降),以及氟达拉滨磷酸盐处理后线粒体NAD⁺水平的变化。后者对于把一个药物的作用机制和NAD⁺转运联系起来是必不可少的数据。
这类课题的一个方法论挑战是:普通比色法试剂盒检测的是整细胞提取物的总NAD⁺,而不是专门的线粒体组分。理论上可以先做线粒体分离再检测,但这在实操中存在明显局限——线粒体分离通常需要15–30 min,全程渗透压和温度的细微变化会让NAD⁺/NADH在分离过程中持续代谢或相互转化,测出来的数值已经不能代表活细胞中的真实状态。
对于SLC25A51这类需要精确区分线粒体内外NAD⁺动态变化的课题,不同方法有各自适合的问题边界:比色法适合整细胞水平的基础表型验证(如敲减前后总NAD⁺含量的组间比较);LC-MS/MS配合同位素示踪可以做代谢流分析,给出各组分的绝对定量;遗传编码荧光探针(如SoNar)则能在活细胞的特定亚细胞器中实时监测NAD⁺/NADH动态,时空分辨率最高,但需要提前构建稳定表达的细胞系。SLC25A51原文发表在Cell Death & Differentiation这个级别,其亚细胞定位的数据支撑需要超出比色法能可靠提供的精度。
比色法在这个方向能做的是:在整细胞层面确认SLC25A51敲减或药物处理前后NAD⁺总量的变化趋势,作为后续更精细实验的预实验或基础表型数据。这是合理的使用场景,但不应被误认为能替代亚细胞分辨率的检测方案。
研究启发
SLC25A51这个方向把NAD⁺研究带进了一个更精细的亚细胞定位问题。类似的逻辑可以延伸到:SIRT1/SIRT3/SIRT5等不同亚细胞定位的sirtuin家族对NAD⁺的竞争性消耗;线粒体功能障碍模型中胞质NAD⁺和线粒体NAD⁺的分别变化;以及其他溶质载体家族成员(SLC25家族其他成员)在代谢重编程中的作用。这些课题的共同前提,都是需要NAD⁺含量的可靠定量作为核心读出。
横向视角:三个方向共同指向的实验逻辑
把这三个课题放在一起看,有一个共同的实验逻辑值得提炼:
NAD⁺/NADH不是一个孤立的氧化还原指标,而是细胞代谢状态的综合读数。 它同时反映了能量代谢效率(TCA循环和呼吸链的运转状态)、信号调控能力(sirtuins的底物可用性)和细胞的应激响应能力(PARP激活后的消耗)。因此,在疾病模型中检测NAD⁺/NADH,相当于同时获取了这几个维度的信息。
三个方向里检测NAD⁺/NADH的具体目的各有侧重:
| 课题方向 | 检测NAD⁺/NADH的核心目的 |
|---|---|
| 慢性肾病(CKD) | 确认病理表型(NAD⁺合成受损的直接证据);建立NAD⁺水平与肾功能指标的相关性 |
| 急性肺损伤(ALI)/HO-1 | 验证保护蛋白的作用机制依赖于NAD⁺(因果链中的关键节点) |
| 肿瘤/SLC25A51 | 证明靶点功能丧失导致线粒体NAD⁺减少(药靶机制验证) |
从样本类型来看,三个方向分别涉及大鼠/小鼠器官组织、培养细胞系和处理后细胞,KTB1020(辅酶Ⅰ NAD(H)检测试剂盒)在这三类样本上均有适用性。组织样本(肾脏、肺组织)约取20 mg,培养细胞约取1×10⁶个,提取流程按各自样本类型的标准操作执行即可。
值得补充的是,文章选取的三个方向并不代表NAD⁺研究的全貌。衰老和神经退行性疾病才是目前NAD⁺研究体量最大、关注度最高的两个领域。NMN(烟酰胺单核苷酸)和NR(烟酰胺核苷)作为NAD⁺前体的补充效果研究,以及老年相关器官功能衰退中NAD⁺水平的系统性下降,是近年来发文量增长最快的方向之一;阿尔茨海默病、帕金森病等神经退行性疾病中,线粒体NAD⁺代谢障碍和SIRT1/SIRT3功能受损之间的联系也在持续被挖掘。
这两个方向同样需要NAD⁺/NADH的定量检测,而且动物组织(脑、骨骼肌、脂肪组织)和体外细胞衰老模型(复制性衰老或药物诱导)都是比色法试剂盒能直接处理的样本类型,不存在亚细胞分辨率的瓶颈问题。衰老领域里顶尖机制研究确实已经大量引入LC-MS和活细胞探针,但在干预效果评估、动物队列的批量组织检测等场景里,比色法仍然是高效且够用的选择。
小结
NAD⁺研究的热度不是偶然的。从慢性肾病到急性肺损伤,从肿瘤增殖到线粒体转运,细胞能量状态的崩塌往往都能在NAD⁺代谢紊乱上找到对应的分子事件。检测NAD⁺/NADH,不是为了凑一个常规生化指标,而是在读一个和细胞命运深度相关的代谢节点。
对于正在设计相关课题的研究者来说,建议在确定检测指标时同时考虑:NAD⁺和NADH的绝对含量(反映合成/消耗的净结果)、NAD⁺/NADH比值(反映氧化还原状态)、以及必要时分亚细胞组分分别检测(区分线粒体和胞质的贡献)。这三个维度的数据组合,能对细胞NAD⁺代谢给出更完整的描述。
CheKine™ KTB1020(辅酶Ⅰ NAD(H)含量检测试剂盒)支持组织、细胞、细菌和血清(浆)四类样本,检测范围0.78–50 µM,可同时定量NAD⁺和NADH并计算比值,适合上述各课题方向的整细胞水平基础检测需求。
参考文献
Li S, et al. Impaired Nicotinamide Adenine Dinucleotide Biosynthesis in the Kidney of Chronic Kidney Disease. Frontiers in Physiology, 2021, 12: 723690. https://doi.org/10.3389/fphys.2021.723690
Yu J, et al. Heme oxygenase-1 protects against endotoxin-induced acute lung injury depends on NAD⁺-mediated mitonuclear communication through PGC1α/PPARγ signaling pathway. Inflammation Research, 2022, 71(9): 1095–1108. https://doi.org/10.1007/s00011-022-01605-y
Luo J, et al. SLC25A51 promotes tumor growth through sustaining mitochondria acetylation homeostasis and proline biogenesis. Cell Death & Differentiation, 2023, 30(8): 1916–1930. https://doi.org/10.1038/s41418-023-01185-2
