白色脂肪褐变研究中的TG甘油三酯检测:为什么必须测,怎么测才准?
前言:脂肪褐变研究的核心矛盾
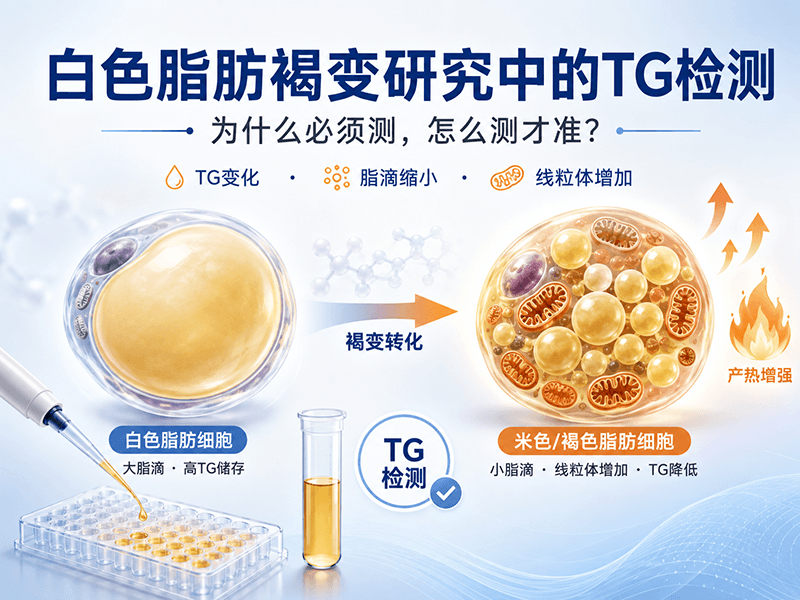
白色脂肪褐变(White-to-beige adipocyte transdifferentiation)是近年代谢研究领域最热门的方向之一。褐色/米色脂肪细胞通过解偶联蛋白1(UCP1)介导的产热机制消耗能量,被认为是对抗肥胖和代谢综合征的潜在靶点。
这个研究方向有一个有趣的内在矛盾:褐变本身就是一个TG消耗的过程。白色脂肪细胞以储存TG为主要功能,而褐变/米色化后,细胞从"储能"模式切换到"耗能"模式——脂滴减小、TG含量下降、线粒体增多、产热基因上调。
因此,TG含量的变化在褐变研究中既是结果指标(褐变成功了吗?),也是机制节点(脂质动员通路是否被激活?)。但正因为如此,TG检测在这个场景下的解读也比MASLD研究更复杂——TG降低不一定意味着褐变成功,也可能只是脂解增加或分化不完全。本文将帮你把这些逻辑理清楚。
一、白色、米色、棕色脂肪的TG代谢差异
理解为什么要测TG,先要理解三种脂肪细胞在TG代谢上的根本差异:
| 脂肪类型 | TG储存量 | 脂滴形态 | 产热能力 | TG动员特征 |
|---|---|---|---|---|
| 白色脂肪(WAT) | 极高 | 单个大脂滴(unilocular) | 无 | 受β肾上腺素受体调控的脂解 |
| 棕色脂肪(BAT) | 低 | 多个小脂滴(multilocular) | 强(UCP1介导) | 冷刺激/交感神经激活后快速动员 |
| 米色脂肪(Beige) | 中等 | 介于两者之间,可向多脂滴转变 | 中等(可诱导) | 类似BAT,受β3-AR激动剂等诱导 |
关键推论:
在褐变研究中,如果你的干预(化合物、基因操控、物理刺激)成功诱导白色脂肪向米色/棕色转化,预期会看到:
- 细胞或组织TG含量下降(脂滴减小)
- 同时伴随UCP1、PGC-1α、CIDEA等产热基因上调
- 线粒体数量和功能指标改善
但TG下降不等于褐变成功——单纯的促脂解处理(如异丙肾上腺素)也会导致TG下降,而不一定伴随真正的褐变表型。这就是为什么TG检测必须与其他指标联合使用,而不能孤立解读。
二、体外模型:3T3-L1脂肪细胞分化中的TG检测
2.1 3T3-L1模型的TG检测时机
3T3-L1前脂肪细胞分化是体外研究脂肪细胞生物学最经典的模型。TG在分化过程中的积累是分化成功的核心标志之一,但在不同阶段检测TG的意义不同:
| 分化阶段 | 检测时机 | 预期TG水平 | 检测目的 |
|---|---|---|---|
| 分化前(Day 0) | 基线 | 极低 | 建立基准值 |
| 分化中期(Day 4–6) | 脂滴开始出现 | 开始升高 | 验证分化启动 |
| 分化完成(Day 8–10) | 脂滴充盈 | 峰值 | 确认分化完全 |
| 褐变诱导后(Day 10+) | 诱导处理后24–72 h | 下降(若褐变成功) | 评估褐变效果 |
⚠️ 重要: 同一批次实验中,对照组(未褐变)和实验组(褐变诱导)必须在同一分化时间点开始处理,否则TG的差异可能来自分化程度不一致而非褐变本身。
2.2 细胞样本TG检测的操作要点
细胞收集:
- 吸去培养基,用预冷PBS轻柔洗涤细胞2次,避免残留培养基中的脂质干扰
- 按实验设计收集细胞:推荐收集5×10⁶个细胞(约2–3个10 cm培养皿的充分分化细胞),计数后记录N
- 800 g离心2 min,弃上清,细胞沉淀置冰上
样本制备:
- 加入1 mL Extraction Buffer,冰浴超声破碎(200W,超声3 s/间隔7 s,重复30次)
- 8000 g,4℃,离心10 min,取上清待测
⚠️ 脂肪细胞的特殊注意事项:
分化成熟的3T3-L1细胞脂滴丰富,普通涡旋或短暂混匀不足以充分破碎脂滴,必须使用超声破碎。超声功率过低(<150W)同样无法有效释放脂滴内的TG,导致结果系统性偏低。
离心后管内会出现三层分层结构,这是脂肪细胞实验中极为常见但容易踩坑的现象:
┌─────────────────────────┐ │ 上层:白色脂饼(Fat Cake)│ ← 脂质密度小于水,浮于最上层 │ 中层:水相上清(目标层) │ ← 含TG提取产物,这才是要吸的 │ 下层:细胞碎片沉淀 │ ← 弃去 └─────────────────────────┘
正确操作: 用移液枪枪头穿过上层脂饼,小心插入中层水相上清,缓慢吸取,避免吸入脂饼或扰动下层沉淀。脂饼越厚(分化越成熟、含脂量越高),这一步越需要耐心。切勿用离心管直接倾倒——脂饼会跟着一起出来。
蛋白定量样本同样存在Fat Cake问题: 另取细胞用去离子水提取蛋白时,离心后同样会出现脂饼,BCA定量时也需要穿过脂饼吸取中层清液,而非直接吸取上层。吸取位置错误是蛋白定量结果异常偏低的最常见原因之一。
脂饼厚度过高(重度脂肪细胞)时,可先在56℃水浴5 min助溶后再离心,有助于减薄脂饼、提高中层上清的可及性。
2.3 细胞TG结果的归一化选择
细胞实验中TG的归一化方式直接影响结论的可靠性,在褐变研究中需要特别谨慎:
按细胞数量归一化(mg/10⁴ cells):适合细胞形态和大小基本一致的比较。但褐变后细胞体积可能发生变化,"相同数量的细胞"代表的生物量不同,可能引入偏差。
按蛋白浓度归一化(nmol/mg prot)——推荐:消除细胞大小和代谢活性差异,是褐变研究中更严谨的归一化方式。同样需要注意:蛋白定量必须另取细胞用去离子水单独提取,并注意穿过脂饼吸取中层清液,不能用TG提取上清。
按DNA含量归一化:在细胞增殖差异较大时,用DNA含量作为分母是最严格的归一化方式,但操作相对复杂,通常用于严格的方法学对照而非常规检测。
前沿方向:mtDNA/nDNA比值归一化:这是近年高分褐变研究中逐渐出现的更精细归一化思路。褐变最核心的细胞学事件是线粒体生物发生(Mitochondrial Biogenesis),褐变成功的细胞线粒体数量显著增加。用实时定量PCR检测线粒体DNA(mtDNA)与核DNA(nDNA)的拷贝数比值(mt/n ratio),不仅可以作为TG数据的归一化分母,本身也是褐变程度的独立定量指标——比UCP1蛋白表达更能直接反映"线粒体真的多了"这件事。
常用的mtDNA引物靶标:mt-CO1(线粒体细胞色素氧化酶1)或 mt-Nd1;nDNA引物靶标:Rpl13a 或 Hk2。这一指标对样本量要求不高,可与TG检测并行从同一批细胞中获取,实验成本增加有限,但数据的深度和前沿性显著提升。
三、体内模型:脂肪组织TG检测的关键差异
体内褐变研究通常涉及皮下白色脂肪组织(scWAT)、附睾白色脂肪(eWAT)和肩胛间棕色脂肪(iBAT)的比较检测,三者在TG水平和操作注意事项上存在显著差异。
3.1 不同脂肪组织的TG水平差异
| 脂肪组织 | 正常小鼠TG参考范围 | 取材特点 | 注意事项 |
|---|---|---|---|
| 皮下WAT(scWAT) | 200–400 mg/g | 质地松软,脂滴丰富 | 最容易诱导褐变,是褐变研究首选 |
| 附睾WAT(eWAT) | 250–450 mg/g | 质地较硬,血管丰富 | 褐变能力弱,更多用作白色脂肪对照 |
| 肩胛间BAT(iBAT) | 80–150 mg/g | 颜色较深,血管密集 | TG本身含量低,冷刺激后进一步下降 |
⚠️ 取材注意: 脂肪组织中含有大量结缔组织、血管和基质细胞,称重前需仔细去除可见的非脂肪组织(筋膜、血管束),否则会稀释TG信号。同一实验批次的取材操作标准必须完全一致。
3.2 冷刺激/β3-AR激动剂处理后的取材时机
冷暴露(4℃,1–7天)或β3-肾上腺素受体激动剂(CL316,243)处理是最常用的体内褐变诱导方案。处理后脂肪组织TG水平会随时间变化,取材时机对结果影响很大:
- 急性冷暴露(24–48 h): BAT的TG快速消耗(下降可达40–60%),scWAT变化较慢
- 慢性冷暴露(7天以上): scWAT开始出现明显褐变表型,TG下降幅度加大,UCP1阳性细胞比例增加
- CL316,243处理(7天): scWAT TG可下降30–50%,同时伴随脂滴多室化(multilocularity)
建议: 取材时间点应与文献中相同干预方案的时间点一致,并在Materials & Methods中明确说明处理时长、剂量和取材时间。
3.3 褐变研究中禁食状态的特殊考量
与肝脏TG检测类似,脂肪组织TG检测同样需要统一禁食状态。但褐变研究有一个额外的复杂因素:禁食本身就是一种促脂解刺激,会导致脂肪组织TG下降,在行为学上类似于冷刺激诱导的产热。
建议: 褐变研究取材前统一禁食4–6小时,并在实验设计中避免将禁食时间与冷刺激/药物处理时间点重叠,防止两种脂解刺激叠加混淆。
四、从文献看TG检测在褐变研究中的典型用法
案例一:褪黑素诱导白色脂肪褐变
发表于 Molecular and Cellular Endocrinology(2024)的研究发现,褪黑素通过其受体MT1介导直接的褐变效应,同时通过诱导M2型巨噬细胞极化间接促进脂肪褐变(Melatonin induces white-to-beige adipocyte transdifferentiation through melatonin receptor 1-mediated direct browning and indirect M2 polarization)。该研究将脂肪细胞TG含量作为褐变程度的功能性验证指标,与UCP1蛋白表达、线粒体膜电位等指标联合构建了完整的褐变表型评估体系。
这个研究的设计提示了一个重要逻辑:TG下降是必要条件,但不是充分条件。只有TG下降同时伴随UCP1上调、产热基因表达增加、线粒体功能改善,才能定义为真正的褐变。
案例二:miR-203的抗肥胖机制
发表于 iScience(2022)的研究揭示了miR-203通过靶向顶端钠依赖性胆汁酸转运体(ASBT)发挥抗肥胖作用(MiR-203 is an anti-obese microRNA by targeting apical sodium-dependent bile acid transporter)。研究者在脂肪组织层面检测了TG含量变化,与血清脂质谱和能量代谢指标联合,证明了miR-203对脂肪储存的系统性影响,而不仅仅局限于肠道。
这个案例的价值在于展示了TG检测在**非典型研究方向(miRNA-脂质代谢轴)**中的桥接作用——当你的研究对象不是经典的脂肪代谢分子时,TG定量数据能把分子层面的调控变化锚定到具体的代谢表型上。
五、褐变研究中的联合指标体系
基础验证组合(确认褐变表型):
- 脂肪细胞/组织TG含量(脂滴消耗的定量指标)
- UCP1蛋白表达(Western blot或免疫组化)
- 产热基因mRNA(UCP1、PGC-1α、CIDEA、PRDM16)
- 脂滴形态(油红O染色,观察单室→多室变化)
扩展组合(机制验证):
- 线粒体功能(Seahorse线粒体呼吸检测,OCR/ECAR)
- 游离甘油释放量(脂解活性的直接指标)
- β氧化相关酶活(CPT1活性)
- 细胞产热(荧光素酶报告基因或TMRE线粒体膜电位)
体内实验追加指标:
- 全身能量代谢(代谢笼,VO₂/VCO₂/RQ)
- 体温变化(冷刺激实验中直肠温度)
- 脂肪组织免疫细胞浸润(流式或免疫组化,M2巨噬细胞比例)
游离甘油干扰的特殊重要性
在褐变研究中,游离甘油干扰比MASLD研究中更需要重视。原因在于:褐变诱导剂(β3-AR激动剂、cAMP激活剂等)本身就是强效脂解刺激,会大幅提升细胞内游离甘油水平。如果不扣除游离甘油背景,测得的"TG降低"有相当一部分可能来自游离甘油背景的升高拉高了空白信号,而非真实TG含量的变化。
操作建议: 在使用任何促脂解类诱导剂的褐变实验中,务必设置游离甘油空白孔(不含脂蛋白酯酶的对照),从总信号中扣除游离甘油背景后再计算TG含量。
六、数据解读:TG变化的三种情景
拿到褐变实验的TG数据后,以下三种情景需要区别对待:
情景一:TG显著下降 + UCP1上调 + 线粒体功能改善→ 典型的成功褐变表型,TG降低来自脂质动员用于产热消耗,结论可靠。
情景二:TG显著下降 + UCP1无变化 + 游离甘油大量释放→ 单纯促脂解效应,不是真正的褐变。检查干预手段是否只激活了脂解通路而未诱导产热基因表达。
情景三:TG无显著变化 + UCP1上调→ 可能的解释:①褐变诱导同时激活了TG合成补偿;②取材时间点过早,脂滴消耗尚未达到统计显著;③样本量不足,效能不够。建议延长处理时间或增加样本量后重复。
总结
在白色脂肪褐变研究中,TG检测是表型验证体系中不可或缺的定量支柱。无论是体外3T3-L1分化模型还是体内脂肪组织取材,TG含量的变化都需要在正确的时间点检测、用合适的归一化方式表达、并与产热基因和线粒体功能指标联合解读,才能得出可靠的结论。
特别需要注意的是,促脂解类诱导剂带来的游离甘油干扰在褐变研究中比其他场景更为突出,游离甘油空白扣除是这个研究方向中的必要步骤,而非可选项。
推荐使用 CheKine™ 甘油三酯(TG)检测试剂盒(微量法,KTB2200) 进行上述实验,灵敏度0.043 mg/mL,可满足从分化早期低TG细胞到充分分化脂肪细胞的全程检测需求,适用于细胞、脂肪组织等多种样本类型。已被发表于 Molecular and Cellular Endocrinology、iScience 等期刊的脂肪褐变与肥胖研究引用验证。
技术咨询:400-6800-830
本文内容仅供科学研究参考,产品不适用于临床诊断。
